Введение
Менее жестко запрограммированная эволюция
Выводы
Осложнения
Хромосомные и химические основы наследственности
Связь классической и молекулярной генетики
Неодарвинизм и популяционная генетика
Холдейн, Фишер и значение отбора
Сьюэлл Райт, адаптивные ландшафты и случай
Рекомендуемая литература
Меланизм у бабочек — подход a posteriori
Приложение теории игр к поведению животных
Изменчивость в природных популяциях; ее значение для адаптации
Адаптации на благо группы и адаптации групп
Адаптации самого полового размножения как такового и порожденные им адаптации
Рекомендуемая литература
Канализация развития
Заключения
Кладизм (5, 7)
Эволюционная систематика (13)
Как образуются таксоны — неодарвинистская точка зрения, или синтетическая теория эволюции
Альтернативная точка зрения — прерывистая эволюция
Сравнение теорий постепенной и прерывистой эволюции
Заключения
Навигация
Эволюционная систематика (13)
Принципы эволюции
242666
знаков
12
таблиц
37
изображений
3. Эволюционная систематика (13)
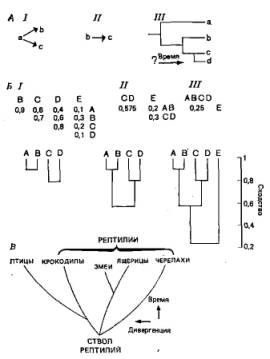
Рис. 5.1. А. Кладисты обычно исходят из допущения, что рассматриваемые виды связаны между собой так, как это показано на схеме /, но не так, как на схеме //. На схеме III показана кладограмма, основанная на последовательных ветвлениях. Вопросительный знак у оси времени стоит потому, что кладограммы можно интерпретировать по крайней мере двумя способами. Согласно одной интерпретации, они представляют собой эволюционные древеса, на которых предки точно не указаны, а поэтому вводится время (классическая кладистика); согласно другой — они изображают типы родственных взаимоотношений, а точки ветвления соответствуют синапо-морфным признакам, так что время не вводится (трансформированная кладистика). Классическую кладистику интересуют эволюционные процессы, а трансформированную — таксономические картины. Конечно, те и другие должны быть как-то связаны между собой, однако некоторые кладисты предпочитают не высказываться относительно точного характера этой связи. Б. Ступенчатое построение фенограммы. Буквы обозначают виды. Числа в матрицах — коэффициенты общего сходства (в диапазоне от 0 — отсутствие сходства до 1 — полное сходство), выведенные на основании всех измеренных признаков. На ступени / матрица содержит коэффициенты сходства для пяти видов. Построение фенограммы начинают с поиска основных пар (для каждого члена такой пары коэффициенты сходства максимальны). Эти основные пары соединяют на соответствующих уровнях оси, по которой отложены коэффициенты сходства. На ступени // каждая основная пара рассматривается как одна единица, и коэффициенты сходства здесь представляют собой средние по всем отдельным видам, то есть 0,575= (0,6+0,4+ + 0,7+0,6)/4. Здесь снова следует найти основные пары и произвести соответствующие соединения на схеме. На ступени /// продолжают и завершают эту процедуру (по Luria S. Е. et al., A. View of Life, Benjamin, 1981). В. Филогенетическое древо, в котором сделана попытка учесть как последовательность ветвления (а тем самым сроки), так и степень дивергенции. Время обычно пытаются отложить как можно точнее на вертикальной оси; горизонтальная ось отражает дивергенцию обычно на основании субъективных оценок, а поэтому довольно неточно.
В эволюционной систематике классификация основана на сочетаний генеалогии и степени общего сходства и различия, отражающего уровень дивергенции. Никаких объективных правил для этого не существует, и принятие того или иного решения в значительной мере зависит от опыта систематиков в оценке относительной значимости различных критериев. Полученные результаты представляют в виде филогенетических древес (рис. 5.1.В).
Некоторые из различий, существующих между этими школами, можно проиллюстрировать на следующих примерах, относящихся к птицам и рептилиям. Как птицы, так и крокодилы происходят от общей предковой группы архозавров, в которую входили динозавры. Какая-то еще более древняя рептилия была общим предком черепах, ящериц и змей, крокодилов и птиц. Поэтому у птиц и крокодилов больше общих признаков, чем у крокодилов и других ныне существующих рептилий.
Однако после того, как птицы отделились от общего ствола, они развивались быстрее, чем крокодилы, и теперь они обладают рядом чрезвычайно своеобразных признаков — способностью к полету, перьевым покровом, гомойотермностью и т. п. Поскольку птицы сильнее дивергировали от предкового ствола, чеш крокодилы, эволюционные систематики отделяют их от всех других рептилий и помещают в отдельный класс — Aves, a всех остальных ныне существующих рептилий, в том числе крокодилов, объединяют в класс Reptilia (рис. 5.1,Б). Фенетики строят аналогичную систему на основе морфологического сходства. Что же касается кладистов, то они при построении систем классификации строго придерживаются генеалогического родства, и поэтому в их кладограммах птицы связаны с крокодилами более тесным родством, чем крокодилы — с черепахами, змеями и ящерицами.
Итак, подводя итоги, можно сказать, что какого-то оптимального способа построения биологической системы классификации, которая отражала бы эволюционные связи, по-видимому, не существует. Кладисты строят ее объективно на основе одной лишь генеалогии, но пренебрегают важными сведениями о дивергенции. Фенетики строят ее объективно, основываясь на сходствах, но неизбежно путают при этом гомологичные и конвергентные признаки. Представители эволюционной систематики пытаются учитывать одновременно и генеалогию, и дивергенцию, но не могут делать это вполне объективно.
Однако независимо от того, какой мы придерживаемся методологии, производя отбор групп однородных таксонов, следует найти их место в линнеевской иерархии, то есть возвести их в определенный ранг, как, например, отряд или семейство. Кладисты часто вводят новый ранг после каждой точки ветвления в кла-дограмме. Эволюционные систематики, однако, судят о ранге таксонов по степени их дивергенции от общего предка, нередко приписывая разные ранги сестринским группам. Определение ранга — процесс крайне субъективный, а поэтому он вызывает многочисленные разногласия и подвержен всевозможным изменениям и уточнениям.
Похожие работы
... матери требовали максимальной "притирки" плода к условиям организма матери, поэтому генетический контроль должен был быть минимальным. 2. Рост мозга. Развитие коры головного мозга. Развитие интеллекта. Эмоциональное развитие. 2.1 Функциональные блоки мозга Дифференциация систем мозговой коры происходит постепенно, и это приводит к неравномерному созреванию отдельных мозговых структур, ...



... . Кроме того, ДНК имеется в митохондриях, которые передаются только по материнской линии. Исследования мутаций в этих ДНК позволяют реконструировать историю биологического развития человечества, происхождение отдельных рас и народностей. Мутация может быть рецессивной, доминантной и полудоминантной в зависимости от состояния гена, в котором она произошла. Гены мутируют с определенной частотой, и ...
— концепция «2К» (кооперация и коммуникация) — эта концепция была разработана в 1900 г. русским ученым-философом П.А.Кропоткиным (1842—1921). В ней идет речь в основном о механизме биологической эволюции. . Термин «эволюция» (лат. evolutio — развертывание) в науке о живом представляет фундаментальное понятие для объяснения возникновения и развития всего живого. Эволюция подразумевает необратимое ...
... о противоречивом пути развития синтетической теории эволюции, о преодолении кризисов в научном познании, направленном на более полное и глубокое понимание закономерностей развития органического мира. Важно отметить, что основы научной теории эволюции заложил Ч. Дарвин. Как господствующее эволюционное учение дарвинизм существовал с 1859 до 1900 гг., т.е. до переоткрытия законов Г. Менделя. До ...
0 комментариев