Введение
Менее жестко запрограммированная эволюция
Выводы
Осложнения
Хромосомные и химические основы наследственности
Связь классической и молекулярной генетики
Неодарвинизм и популяционная генетика
Холдейн, Фишер и значение отбора
Сьюэлл Райт, адаптивные ландшафты и случай
Рекомендуемая литература
Меланизм у бабочек — подход a posteriori
Приложение теории игр к поведению животных
Изменчивость в природных популяциях; ее значение для адаптации
Адаптации на благо группы и адаптации групп
Адаптации самого полового размножения как такового и порожденные им адаптации
Рекомендуемая литература
Канализация развития
Заключения
Кладизм (5, 7)
Эволюционная систематика (13)
Как образуются таксоны — неодарвинистская точка зрения, или синтетическая теория эволюции
Альтернативная точка зрения — прерывистая эволюция
Сравнение теорий постепенной и прерывистой эволюции
Заключения
Навигация
Сравнение теорий постепенной и прерывистой эволюции
Принципы эволюции
242666
знаков
12
таблиц
37
изображений
5.5. Сравнение теорий постепенной и прерывистой эволюции.
Для того чтобы показать более конкретно различие между постепенной и прерывистой эволюцией в плане эволюционных направлений и адаптации, рассмотрим классический пример эволюции современной лошади, далеким предком которой был Hyracotherium — трехпалое существо величиной не больше собаки. Палеонтологическая летопись свидетельствует, по-видимому, о постепенном изменении размеров тела и формы, приведшем в конечном счете к хорошо знакомому нам Equus. Классическое объяснение с позиций постепенной эволюции сводится к прогрессивному изменению под действием естественного отбора (см. разд. 3.2). В противоположность этому теория прерывистого равновесия объясняет направления в эволюции лошадей как результат асимметрии генеалогического древа. Эволюционная история лошади рассматривается как видообразовательная линия, в которой возникали некоторые новые виды, отличавшиеся большими размерами тела и сокращением числа пальцев, а также другие виды с совершенно иными морфологическими изменениями. Виды с более крупным телом и редуцированными пальцами оказались более удачливыми, чем виды с более примитивными признаками — большим числом пальцев и меньшими общими размерами, — и это привело к асимметрии генеалогического древа. С течением времени «центр тяжести» явно склоняется в сторону более крупного вида с одним пальцем (как, например, на рис. 5.4).
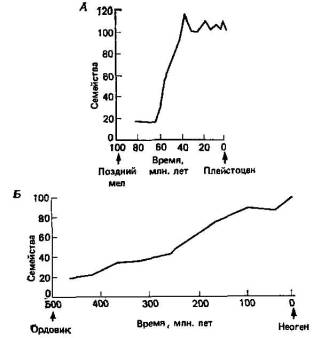
Рис. 5.5. Разнообразие семейств млекопитающих (А) и двустворчатых моллюсков (Б) на протяжении длительных периодов времени (19).
В табл. 5.2 перечислены главные различия между теориями постепенной и прерывистой эволюции. Таблица 5.2. Сравнение гипотез постепенного и прерывистого видообразования
| Фактор | Постепенное видообразование | Прерывистое видообразование |
| Изоляция | Случайный процесс, иногда неслучайный обычно аллопатрический | Изолированная подгруппа часто имеет неслучайный генетический состав; может быть симпатрическим или парапат-рическим |
| Источник изменчивости | Главным образом точковые мутации | Главным образом макромутации (то есть мутации в первоначальном смысле) |
| Отбор | Внутри популяций | Между популяциями (или видами) |
| Дивергенция | Медленная и непрерывная (и пропорциональная числу поколений) | Быстрая (непропорциональная числу поколений), но при отсутствии изменений в течение длительных периодов |
| Ограничения | Не очень существенные | Очень существенные |
| Адаптации | Очевидные и совершенные | Не столь очевидные и менее совершенные |
5.6. Проверка двух теорий эволюции.
Стенли (18,19) рассмотрел разного рода данные, которые можно использовать для сравнительной оценки теорий постепенной и прерывистой эволюции. К наиболее общим соображениям относится ожидание, что скорость макроэволюции (то есть скорость возникновения семейств и отрядов, отражающих крупные морфологические изменения) должна быть: 1) согласно модели постепенной эволюции, пропорциональна времени или, точнее, числу поколений, на протяжении которых могло развиваться разнообразие; и 2) согласно модели прерывистой эволюции, пропорциональна степени расщепления, то есть видообразования, которое имело место. Если удастся оценить как время, так и скорость расщепления, то тогда мы в принципе сможем провести различие между этими двумя гипотезами. На этой основе Стенли проводит сравнение между млекопитающими и двустворчатыми моллюсками. У млекопитающих скорость видообразования высокая и радиация в пределах группы, в результате которой возникло примерно 100 семейств, происходила в течение менее чем 30 млн. лет. У двустворчатых моллюсков, напротив, видообразование протекало медленно и соответственно происходила медленная радиация на протяжении более чем 300 млн. лет (рис. 5.5). Более того, среди двустворчатых моллюсков у одной группы сердцевидок (сем. Cardiidae) так называемого понтического ряда, изолировавшегося из Средиземноморья в Каспийском море, видообразование происходило быстрее, чем у ее близких родичей, и за менее чем 3 млн. лет эта группа достигла такого морфологического разнообразия, что ее теперь делят на четыре подсемейства.
Следствие, вытекающее из всех этих рассуждений, состоит в том, что в группах, в которых на протяжении длительных периодов геологического времени происходило мало эволюционных изменений, видообразование, очевидно, было незначительным. И действительно, так, по-видимому, обстояло дело со многими так называемыми живыми ископаемыми. Хорошим примером служат двоякодышащие рыбы. Быстрые морфологические изменения происходили у них только в период быстрого видообразования, в середине палеозоя, вскоре после их появления. После этого скорость видообразования резко понизилась, и вместе с ней упала и скорость морфологических изменений. Другим примером служат аллигаторы, строение которых в процессе эволюции изменялось мало, которые представлены в настоящее время всего двумя видами и никогда не отличались большим числом видов. Наконец, известен лишь один ныне живущий вид трубкозубов, а в палеонтологической летописи, вплоть до начала миоцена, к которому относятся самые ранние находки представителей этой группы, обнаружено лишь очень небольшое число их линий.
Все это свидетельствует в пользу теории прерывистой эволюции. Однако имеющиеся данные нельзя считать решающими, так как их анализ зависит главным образом от разделения ископаемых таксонов, которое может отражать в большей мере пристрастия систематиков, чем истинное течение эволюции; так, например, вопрос о том, какие группы видов образуют семейства, довольно субъективен. Даже соглашаясь принимать фактические данные, классический неодарвинист может заявить, что ряд факторов и, в частности, вариации в интенсивности естественного отбора перекрывают ожидаемую корреляцию между скоростью микроэволюции и временем, предсказываемую в пункте 1 в начале этого параграфа. Иными словами, неодарвинизм не настаивает на непременной постепенности. Он допускает возможность периодов быстрых изменений под действием интенсивного отбора и периодов медленных изменений под действием слабого отбора. Кроме того, застой в морфологии твердых частей тела — а именно они сохраняются в палеонтологической летописи—не обязательно означает застой в строении мягких частей, а также метаболических и поведенческих процессов. Таким образом, хотя фактические данные, очевидно, свидетельствуют в пользу теории прерывистой эволюции, они все еще далеко не окончательны, и вопрос остается открытым, вызывая горячие дебаты.
5.7. Была ли эволюция человека постепенной или прерывистой?
Креационисты (см. гл. 1) считали, что человек отделился от человекообразных обезьян с того момента, когда бог вдохнул в него душу! В ответ на это дарвинисты и неодарвинисты упорно настаивают на том, что человек произошел от человекообразных обезьян в результате постепенной прогрессивной эволюции. Исходя из этого, обычно принято помещать вид Homo sapiens в конец непрерывного ряда, после по крайней мере двух других видов (как, например, на рис. 5.6). Эти разные виды характеризуются следующим образом.
1. Ramapithecus (от Рама —индийское божество, и греч. пи-текос —обезьяна). Эта группа существовала примерно 8— 14 млн. лет назад. Она известна только по фрагментам нижней челюсти, которые позволяют считать, что лицевой отдел черепа у рамапитека был плоским, со слабо выступающей нижней
частью.
2. Австралопитек (от лат. austral —южный). Эта группа существовала 1,5—5 млн. лет назад. Лицевой отдел черепа у австралопитеков был плоским. Расположение черепа относительно позвоночника и костей задних конечностей относительно таза позволяет считать, что положение тела у него было вертикальным. Известны две главные группы австралопитеков: Australopithecus robustus с массивным черепом и крупными зубами, что свидетельствует о растительной диете, и группа, к которой относятся A. afarensis и A. africanus, с более изящным скелетом.
3. Homo habilis (человек умелый). Эта группа существовала 1,5—2 млн. лет назад. Ее представители были прямоходящими, имели относительно большой головной мозг и пользовались различными орудиями.
4. Homo erectus (человек прямоходящий). Эта группа существовала 1 млн. лет назад. У ее представителей походка была еще более сходна с нашей, мозг был крупнее, чем у Я. habilis, и они пользовались огнем.
5. Неандертальцы (от названия долины Неандерталь, где они были впервые найдены). Неандертальцы существовали примерно 0,5 млн. лет назад; они настолько сходны с современными людьми как по морфологии, так и по наличию определенной культуры с различными обрядами, что их считают одним из подвидов Homo sapiens.
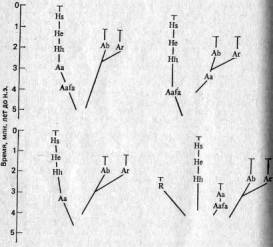
Рис. 5.6. Две разные филогении, предложенные для основных таксонов гоминид (2). R — Ramapi-thecus; Aafa—Australopithecus afarensis; Aa — A. africanus; Ab—A. boisei; Ar — A. robustus; Hh — Homo habilis; He — H. erectus; Hs — H. sapiens.
Недавно были обнаружены два важных факта, которые возродили точку зрения о том, что эволюция человека — процесс прерывистый (19). Во-первых, было высказано мнение, что одновременно существовал не один, а несколько видов гоминид, а это указывает на то, что филогения рода Homo была кустистой, а не линейной. Во-вторых, полагают, что имеются данные, свидетельствующие о наличии относительно длительных периодов застоя в филогении как группы автралопитеков, так и гоминид (см., например, Australopithecus africanus, Homo habilis и Я. erectus на рис. 5.7), а это довольно удивительно, принимая во внимание высокую абсолютную скорость, характерную для эволюции человека.
Вместе с тем данные о перекрывании во времени между видами рода Homo не бесспорны, и если даже они верны, то не отвергают интерпретацию эволюции человека как процесса постепенного.
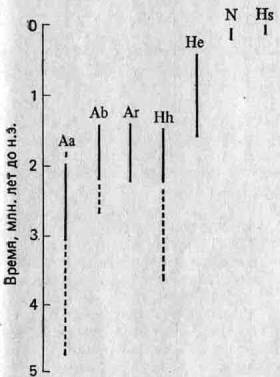
Рис. 5.7. Стратиграфические диапазоны гоминид (19). Прерывистыми линиями обозначены ископаемые остатки, предположительно отнесенные к тому или иному виду. Обозначения те же, что на рис. 5.6. N — неандерталец.
Ветвление могло просто увеличить число направлений, по которым эволюция человека протекала постепенно. Кроме того, что гораздо важнее, хотя разные таксоны гоминид резко различаются по некоторым морфологическим признакам, например по признакам черепа, количественные признаки, например размеры тела и емкость черепной коробки, изменяются почти непрерывно (рис. 5.8). Кроме того, по мнению некоторых антропологов, застой в пределах отдельных таксонов скорее кажущийся, чем реальный (2). Например, у самых ранних экземпляров Н. erectus лицевой отдел черепа более вытянутый и плоский, черепная коробка ниже, тоньше, менее прочна и емкость ее меньше, чем у более поздних Н. erectus.
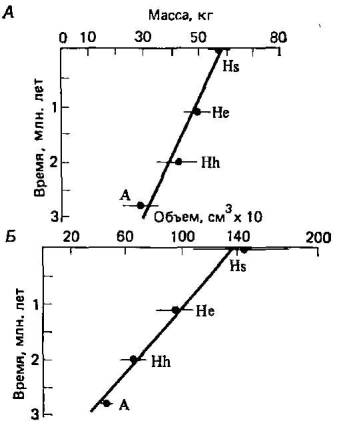
Рис. 5.8. Изменения среднего веса тела (Л) и объема черепной коробки (Б) у гоминид с течением времени (2). Обозначения те же, что на рис. 5.6.
Похожие работы
... матери требовали максимальной "притирки" плода к условиям организма матери, поэтому генетический контроль должен был быть минимальным. 2. Рост мозга. Развитие коры головного мозга. Развитие интеллекта. Эмоциональное развитие. 2.1 Функциональные блоки мозга Дифференциация систем мозговой коры происходит постепенно, и это приводит к неравномерному созреванию отдельных мозговых структур, ...



... . Кроме того, ДНК имеется в митохондриях, которые передаются только по материнской линии. Исследования мутаций в этих ДНК позволяют реконструировать историю биологического развития человечества, происхождение отдельных рас и народностей. Мутация может быть рецессивной, доминантной и полудоминантной в зависимости от состояния гена, в котором она произошла. Гены мутируют с определенной частотой, и ...
— концепция «2К» (кооперация и коммуникация) — эта концепция была разработана в 1900 г. русским ученым-философом П.А.Кропоткиным (1842—1921). В ней идет речь в основном о механизме биологической эволюции. . Термин «эволюция» (лат. evolutio — развертывание) в науке о живом представляет фундаментальное понятие для объяснения возникновения и развития всего живого. Эволюция подразумевает необратимое ...
... о противоречивом пути развития синтетической теории эволюции, о преодолении кризисов в научном познании, направленном на более полное и глубокое понимание закономерностей развития органического мира. Важно отметить, что основы научной теории эволюции заложил Ч. Дарвин. Как господствующее эволюционное учение дарвинизм существовал с 1859 до 1900 гг., т.е. до переоткрытия законов Г. Менделя. До ...
0 комментариев