Введение
Менее жестко запрограммированная эволюция
Выводы
Осложнения
Хромосомные и химические основы наследственности
Связь классической и молекулярной генетики
Неодарвинизм и популяционная генетика
Холдейн, Фишер и значение отбора
Сьюэлл Райт, адаптивные ландшафты и случай
Рекомендуемая литература
Меланизм у бабочек — подход a posteriori
Приложение теории игр к поведению животных
Изменчивость в природных популяциях; ее значение для адаптации
Адаптации на благо группы и адаптации групп
Адаптации самого полового размножения как такового и порожденные им адаптации
Рекомендуемая литература
Канализация развития
Заключения
Кладизм (5, 7)
Эволюционная систематика (13)
Как образуются таксоны — неодарвинистская точка зрения, или синтетическая теория эволюции
Альтернативная точка зрения — прерывистая эволюция
Сравнение теорий постепенной и прерывистой эволюции
Заключения
Навигация
Рекомендуемая литература
Принципы эволюции
242666
знаков
12
таблиц
37
изображений
3.9. Рекомендуемая литература.
Критическое рассмотрение адаптационистской программы см. у Гоулда и Левонтина (18). Обоснование этой программы дает Доукинс (13). Использование принципа оптимальности иллюстрирует Александер (1), а теории игр —Мэйнард Смит (29). Приложение адаптационистской программы к физиологической экологии рассматривают Таунсенд и Кейлоу (40), а к экологии поведения— Кребс и Девис (23).
Глава 4. ЭВОЛЮЦИЯ И РАЗВИТИЕ.
4.1. Введение.
При неодарвинистском подходе к эволюционной биологии главное внимание уделяется генетическим основам изменения и постоянства популяций, тогда как адаптационисты сосредоточили внимание на фенотипах. Однако эти два аспекта эволюции разделять не следует, ибо выражение фенотипа частично зависит от генотипа, а распространение генов зависит от того, сколь успешно фенотипы взаимодействуют со своей средой. Эти взаимозависимости иллюстрирует рис. 4.1. Экспрессия генов осуществляется через посредство морфогенетической системы, на которую оказывают также влияние изменяющиеся факторы среды. Уоддингтон (12) назвал изучение этих взаимодействий эпи-генетикой. В гл. 3 мы уже коснулись экологических и реверти-рованных эпигенетических законов, показанных на рис. 4.1.
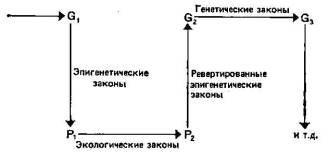
Рис. 4.1. Некое исходное распределение генов (Gi) дает определенные фенотипы (Pi) в соответствии с эпигенетическими законами. Распределение фенотипов изменяется в соответствии с экологическими законами (переходя в pz)- Эти фенотипы вносят в генофонд (Gj) гены, которые распределяются в соответствии с генетическими законами (Оз). В этой главе основное внимание уделено рассмотрению пути от генов (G) к фенотипам (Р).
Здесь мы обратимся к самим эпигенетическим законам. Будут рассмотрены лишь два из наиболее важных принципов: как мелкие мутации могут амплифицироваться в процессе развития и как морфогенетическая система противодействует некоторым типам генетических изменений.
4.2. Амплификация в процессе развития.
Джозеф Нидхем (9) различал в онтогенезе три основных процесса: 1) рост — увеличение объема и веса; 2) развитие — диф-ференцировка и возрастание сложности организации (морфогенез); 3) созревание — прекращение развития и роста и наступление репродуктивных процессов. Джулиан Хаксли (7) изучал, каким образом второй из этих процессов взаимодействует с первым, направляя и регулируя изменения размеров органов и соотнося их друг с другом и с изменениями величины организма в целом. Такие взаимоотношения часто описываются простым математическим уравнением
![]()
где У — размеры данной части, X — размеры другой части или организма в целом, а и р — константы (но ср. Смит (11)).
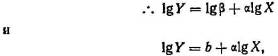
где b=lg(бета). Эти соотношения известны под названием алломет-рических зависимостей, где а — константа аллометрии: если а>1, то У увеличивается в размерах быстрее, чем X, если же <х<1, то наоборот, если а=1, то X и У растут пропорционально одна другой, и этот особый случай называют изометрией. Соответствующие примеры приведены на рис. 4.2. Эти зависимости могут отражать лежащие в их основе морфогенетические ограничения, и (или) скейлинг-эффект, и (или) конкуренцию между отдельными частями организма за ограниченные ресурсы, поступающие с пищей (11).
Нидхем (9) указывал, что как в принципе, так и практически относительные скорости и сроки наступления трех выделенных им фундаментальных процессов можно изменять и что такие регулировки оказывают глубокое влияние на конечный результат развития. Можно, например, экспериментально ускорить дифференцировку по отношению к росту и получить карликов (экспериментальная карликовость). У некоторых беспозвоночных можно подавить размножение и получить великанов (паразиты нередко кастрируют своих хозяев, что приводит к тому же результату). Если возможно искусственно разделить рост и развитие, то можно предполагать, что такая гетерохрония играет важную роль в эволюции. Мелкие мутации могли бы оказывать некоторое влияние на скорости и сроки роста органов и тканей, но сильно воздействовать на конечный результат — иными словами, небольшие изменения в а и (или) b могут иметь очень важные последствия для организации и морфологии взрослого организма.
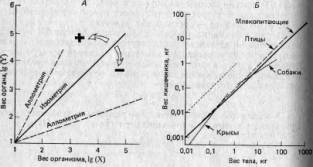
Рис. 4.2. Примеры аллометрии в теории (Л) и на практике (Б). Построенные в логарифмическом масштабе графики зависимости размеров одного органа (Y) от другого или от размеров организма в целом (X) часто имеют вид прямых. Если наклон этих прямых равен единице, то зависимость называют изометрической (для сравнения на рис. Б она изображена пунктиром); в этом случае X и Y увеличиваются в размерах с одинаковыми скоростями. Зависимость между весом кишечника и общим весом тела у млекопитающих и птиц примерно изометрична. Если наклон больше единицы, то зависимость положительно аллометрична и размеры Y увеличиваются быстрее, чем размеры X (например, данные по крысам). Когда наклон меньше единицы, то зависимость отрицательно аллометрична и Y увеличивается медленнее, чем X (например, данные по собакам). Такие графики можно строить для особей, принадлежащих к одному виду (например, данные по собакам и крысам), или для особей, принадлежащих к нескольким разным видам (например, данные по млекопитающим и птицам).
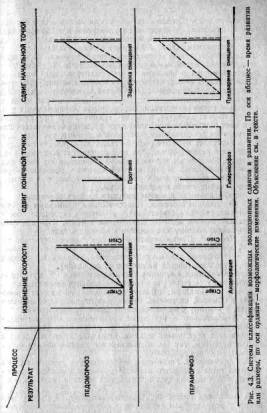
Олберч и др. (2) систематизировали возможные способы возникновения таких регулировок по отношению к некой временной компоненте, определяющей инициацию развития, возраст и начало созревания (то есть 3-й процесс из триады Нидхема). На рис. 4.3 эта система представлена графически. В каждом квадрате заключена некая траектория развития, то есть некий показатель формы в зависимости от размеров или возраста. (Если используются размеры, то эти графики можно рассматривать как эквиваленты зависимостей, которые исследовал Хаксли.) Сплошными линиями изображены траектории предков, а прерывистыми — потомков. Вертикальная линия «Старт» указывает начало развития, а линия «Стоп» — его прекращение. При всех изменениях в верхнем ряду развитие замедляется или урезывается по сравнению с развитием предков, так что зрелость наступает на более ранней стадии развития. Этот процесс называется педоморфозом. При всех изменениях в нижнем ряду развитие, напротив, ускоряется или удлиняется, и это носит название пераморфоза. В этих случаях потомки проходят через предковые стадии на более раннем этапе развития, то есть имеет место рекапитуляция предковых форм.
Можно привести примеры каждого из этих процессов.
1. Ретардация/неотения. Классическими примерами служат некоторые амфибии (хвостатые), у которых взрослые особи сохраняют жабры и другие личиночные органы. Взрослая форма выглядит поэтому как личинка и обитает в воде.
2. Прогенез. Самцы некоторых ракообразных достигают половой зрелости в то время, когда их общие размеры невелики. Эти миниатюрные самцы «паразитируют» на самках, которые гораздо крупнее (иногда на несколько порядков).
3. Предварение смещения (predisplacement). У мутантных «ползающих» (screeper) кур кости ног дифференцируются позднее, чем у нормальных цыплят, но дальнейший их рост происходит с обычной скоростью. Время вылупления также не изменяется, а поэтому у вылупляющихся цыплят ноги короткие.
4. Акселерация. У некоторых сравнительно молодых видов аммонитов рост лопастных линий на раковинах ускорен по сравнению с предковыми видами. Однако в этом замешаны и другие процессы, так что в дальнейшем данный пример будет рассмотрен более подробно.
5. Гиперморфоз. Гигантские по сравнению с другими видами размеры ныне вымершего оленя Megaloceros giganteus достигались, возможно, за счет продолжения роста и задержки размножения.
6. Задержка смещения (postdisplacement). Этот процесс вместе с предварением смещения, возможно, участвует в регуляции типов полосатости у зебры. Бард (3) считает, что у зародыша имеется единственный механизм, создающий вертикальные полосы. В зависимости от того, на какой стадии роста он вступает в действие, полосы на голове будут шире (процесс начинается раньше — предварение смещения) или уже (процесс начинается позднее — задержка смещения) и расположение полос может быть весьма разнообразным. Наблюдается несколько типов полосатости, которые можно объяснить инициацией процесса образования полос в течение третьей (Equus burchelli), четвертой (Е. zebra) или пятой (Е. grevyi) недель развития.
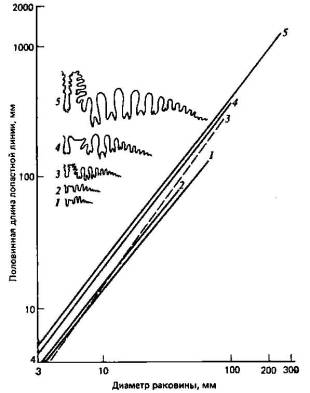
Рис. 4.4. Участие нескольких морфогенетических явлений в эволюции аммонитов (8). Изображены лопастные линии предковых (/) и происходящих от них видов (2—5). Самая древняя часть лопастной линии — небольшой участок справа. Более подробные объяснения см. в тексте.
Не следует, однако, ожидать, что нам всегда удастся наблюдать «в чистом виде» описанные выше изменения; в сущности, более вероятно возникновение морфологических изменений в результате сочетания различных морфогенетических явлений. Хорошим примером служит аллометрическая зависимость между длиной лопастной линии и общими размерами тела у аммонитов, о которой уже говорилось выше и которая представлена на рис. 4.4. Здесь наклон кривых для потомков круче, чем для предков (рис. 4.4,1), так что дифференцировка этих лопастных линий у более молодых видов ускорена по сравнению с предковыми видами. Но, кроме того, в филогенезе этой группы наблюдается выраженное увеличение общих размеров, и у потомков продолжается аллометрический рост лопастных линий, выходящий далеко за пределы размеров взрослых особей у предковых видов (гиперморфоз). Наконец, траектории видов-потомков располагаются выше, чем траектории предковых видов, так что потомки с самого начала уже обладают известным преимуществом, возможно, в результате какого-то предварения смещения.
Итак, все приведенные выше примеры показывают, каким образом изменения процессов развития могут оказывать существенное воздействие на дефинитивную морфологию взрослого организма. В принципе это может быть достигнуто при помощи мелких изменений в скоростях нескольких ключевых процессов, то есть путем мелких генетических изменений. В этом и заключается амплификация в процессе развития.
Похожие работы
... матери требовали максимальной "притирки" плода к условиям организма матери, поэтому генетический контроль должен был быть минимальным. 2. Рост мозга. Развитие коры головного мозга. Развитие интеллекта. Эмоциональное развитие. 2.1 Функциональные блоки мозга Дифференциация систем мозговой коры происходит постепенно, и это приводит к неравномерному созреванию отдельных мозговых структур, ...



... . Кроме того, ДНК имеется в митохондриях, которые передаются только по материнской линии. Исследования мутаций в этих ДНК позволяют реконструировать историю биологического развития человечества, происхождение отдельных рас и народностей. Мутация может быть рецессивной, доминантной и полудоминантной в зависимости от состояния гена, в котором она произошла. Гены мутируют с определенной частотой, и ...
— концепция «2К» (кооперация и коммуникация) — эта концепция была разработана в 1900 г. русским ученым-философом П.А.Кропоткиным (1842—1921). В ней идет речь в основном о механизме биологической эволюции. . Термин «эволюция» (лат. evolutio — развертывание) в науке о живом представляет фундаментальное понятие для объяснения возникновения и развития всего живого. Эволюция подразумевает необратимое ...
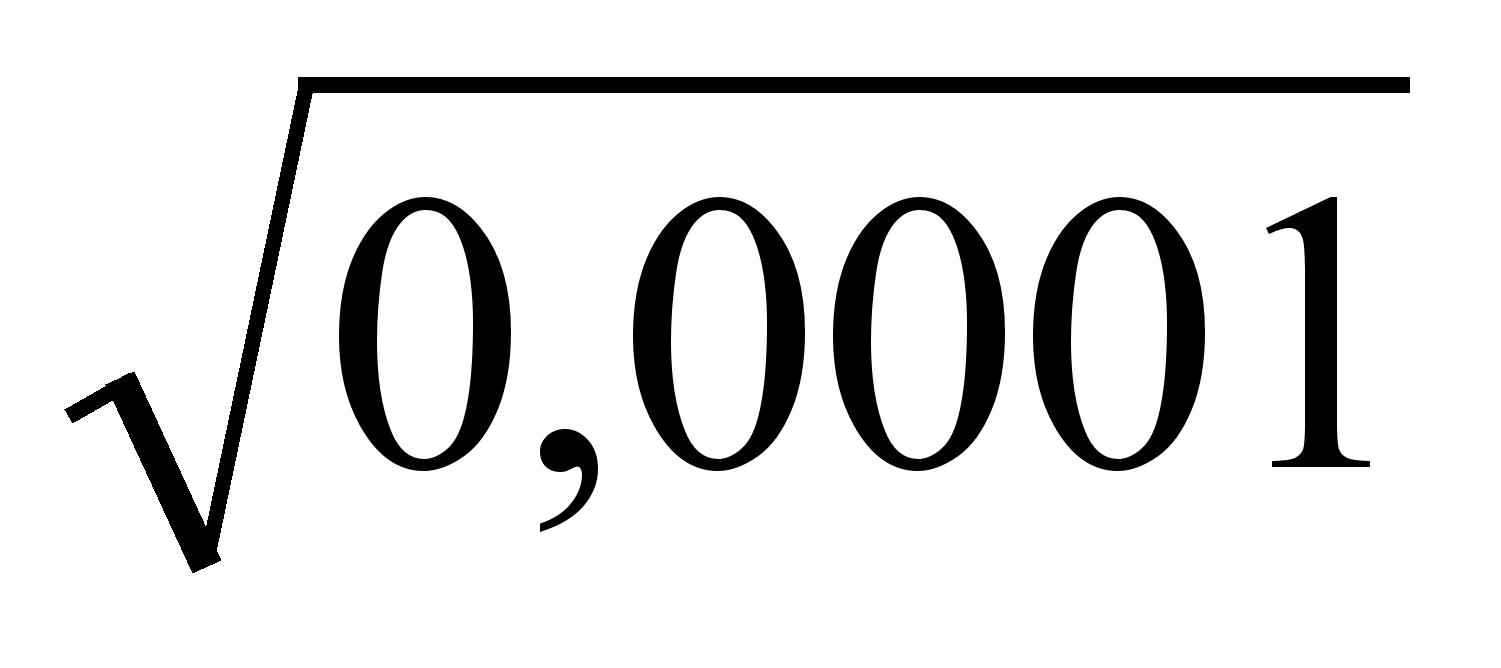
... о противоречивом пути развития синтетической теории эволюции, о преодолении кризисов в научном познании, направленном на более полное и глубокое понимание закономерностей развития органического мира. Важно отметить, что основы научной теории эволюции заложил Ч. Дарвин. Как господствующее эволюционное учение дарвинизм существовал с 1859 до 1900 гг., т.е. до переоткрытия законов Г. Менделя. До ...
0 комментариев