Введение
Менее жестко запрограммированная эволюция
Выводы
Осложнения
Хромосомные и химические основы наследственности
Связь классической и молекулярной генетики
Неодарвинизм и популяционная генетика
Холдейн, Фишер и значение отбора
Сьюэлл Райт, адаптивные ландшафты и случай
Рекомендуемая литература
Меланизм у бабочек — подход a posteriori
Приложение теории игр к поведению животных
Изменчивость в природных популяциях; ее значение для адаптации
Адаптации на благо группы и адаптации групп
Адаптации самого полового размножения как такового и порожденные им адаптации
Рекомендуемая литература
Канализация развития
Заключения
Кладизм (5, 7)
Эволюционная систематика (13)
Как образуются таксоны — неодарвинистская точка зрения, или синтетическая теория эволюции
Альтернативная точка зрения — прерывистая эволюция
Сравнение теорий постепенной и прерывистой эволюции
Заключения
Навигация
Холдейн, Фишер и значение отбора
Принципы эволюции
242666
знаков
12
таблиц
37
изображений
2.3.2. Холдейн, Фишер и значение отбора
Может ли естественный отбор распознавать мелкие различия в генетически детерминированных признаках, оказывающие лишь небольшое воздействие на выживание и размножение их носителей? Могли ли эти мелкие случайные различия сами по себе вызывать существенные изменения? Или же нам следует допустить, что основным материалом для эволюции служат крупные скачкообразные изменения? Эти вопросы поставили Дж. Б. С. Холдейн в своей книге «Генетическая теория эволюции» (J. В. S. Haldane, Genetlcal Theory of Evolution, Longmans, 1930) и Р. А. Фишер в «Факторах эволюции» (R. A. Fisher, The •Causes of Evolution, Oxford, 1932). Эти две книги сыграли чрезвычайно важную формирующую роль в дальнейшем развитии неодарвинизма. Еще одной важной фигурой в его развитии был Сьюэлл Райт (Sewall Wright). Однако, подход Райта к этим вопросам настолько отличался от позиций Холдейна и Фишера, что мы рассмотрим его отдельно в разд. 2.3.3.
И Фишер, и Холдейн выражали интенсивность отбора через изменения в частотах генов. Те аллели, частота которых относительно других аллелей выше, считаются более приспособленными, поэтому изменение относительных частот аллелей служит мерой «неодарвиновской» приспособленности. (Обратите внимание, что нельзя ограничиваться рассмотрением абсолютных изменений, так как при понижении абсолютной плотности данного аллеля в популяции он все же может оставлять больше своих представителей, чем другие аллели, если плотность этих других аллелей уменьшается быстрее. В таком случае он все еще более приспособлен, поскольку в преобладающих в данной среде условиях он по сравнению с другими аллелями оказался более преуспевающим.)
Проще всего начать анализ с динамики генов в гаплоидных популяциях (то есть в популяциях, в которых каждая особь содержит по одному аллелю на локус); в табл. 2.2 представлены основные расчеты для двух аллелей (Л и а) в такой популяции. Холдейн измерял приспособленность, сравнивая чистые скорости прироста R, то есть делил значение R для каждого аллеля на максимальное значение R, так что приспособленность наиболее преуспевающего аллеля становится равной 1, а приспособленность других аллелей — какой-то доле 1. Этот индекс обычно обозначают буквой w и называют селективной или адаптивной ценностью. В некоторых случаях эту величину можно записать в виде 1 —w ( = s), что выражает степень, в которой данный ген подвергается отрицательному отбору; s называют коэффициентом отбора.
Таблица 2.2. Вычисление приспособленности в гаплоидной популяции
| Аллель | Численность в одном поколении | Численность в одновременном следующем поколении | Чистый прирост за период между нулевым временем и временем t | Показатель приспособленности ' Холдейна (относительно Л) |
| А | N0 | Nt | Nt --- = RA N0 | 1 |
| а | N0 | nt | nt ---- = Ra n0 | Ra/RA |
Это крайне упрощенный пример. Время генерации у аллелей А и а может быть различным, и в таком случае придется вносить поправку путем деления всех величин на время генерации. Если поколения перекрываются (то есть особи, составляющие популяцию в данный момент времени, имеют разный возраст) и размножение у разных поколений происходит несинхронно, то положение еще более осложняется, потому что в этом случае нет стандартных интервалов, которые можно было бы использовать для сравнения изменений частоты аллелей А и а. Поэтому Фишер предпочел использовать в качестве меры приспособленности относительные скорости размножения генов в данной популяции, так как в экстремальном случае непрерывного размножения (какие-то особи из тех или иных поколений приносят потомков все время) эту скорость можно вычислить из изменений абсолютной численности (например, N для аллеля А и п для аллеля а) на протяжении какого-либо промежутка времени. Подобный мультипликативный рост описывается уравнениями dN/dt—rAN и dn/dt = ran, которые после интегрирования приобретают вид Nt = N0erAt и п( = поега( (е — основание натуральных логарифмов, r — константа, t — время, А к а — аллели, к которым относится данный параметр). Коэффициент г, который Фишер назвал «мальтузианским параметром» (поскольку Мальтус был одним из первых, создавших модель мультипликативного роста), представляет собой меру скорости распространения каждого гена и поэтому служит показателем приспособленности. Обратите внимание, что если мы примем t за среднее время генерации, как в случае прерывистого роста, то тогда
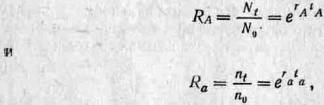
и если, из соображений удобства, мы примем tA = ta=, то RalRA = era~-rAn поэтому In wa=ra—га- Следовательно, эти два показателя приспособленности связаны друг с другом. Популицонные генетики обычно предпочитают пользоваться холдейновской мерой приспособленности, тогда как эволюционные экологи часто используют меру, предложенную Фишером. В этом разделе мы в дальнейшем будем пользоваться параметрами w и s, а к коэффициенту r вернемся в разд. 3.4. (Более подробное рассмотрение коэффициента r и его ограничений в качестве меры приспособленности можно найти у Чарлзуорта (3).)
Уравнения для диплоидного случая гораздо сложнее, чем для гаплоидного, потому что степень распространения или сокращения данного аллеля в популяции зависит от того, с какими аллелями он становится связанным в гетерозиготах. Так, например, вредный рецессивный ген будет защищен от элиминации отбором, если он в гетерозиготном состоянии связан с благоприятными доминантными аллелями. И напротив, отбор, направленный против вредного доминантного аллеля, ведет к элиминации благоприятных рецессивных аллелей, если они связаны с ними в гетерозиготах.
Приспособленность генотипов можно вычислить в основном таким же способом, как и приспособленность составляющих их аллелей. Это довольно искусственный способ, потому что гомо-зиготы одного поколения могут вносить вклад в гетерозиготы другого и наоборот (то есть поставляются последующим поколениям гены, а не генотипы). Тем не менее из этих частот генотипов можно вывести изменение частоты аллелей при условии, что мы располагаем дополнительной информацией: а) о начальных частотах аллелей и б) о степени доминирования. Мы делаем это, используя адаптивные ценности и коэффициенты отбора, а не мальтузианские параметры, однако аналогичным образом можно использовать и последние (9).
Простейший возможный случай относится к одному локусу с двумя аллелями, например Л и а, с частотами соответственно р и q. Большая часть рассуждений в популяционной генетике основана на системе именно такого типа. Допустим, что в результате отбора равновесие Харди—Вайнберга в некоей воображаемой популяции сдвигается следующим образом:
![]()
Изменение, происшедшее с каждым генотипом (то есть эквивалент R в табл. 2.2), составит поэтому АА = 0,32/0,16 = 2; Аа — = 0,52/0,48=1,08; аа = 0,16/0,36=0,44; на основе этого можно получить значение приспособленности для каждого генотипа, разделив каждое из полученных чисел на 2 (то есть на самое большое значение).
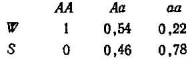
Отсюда (W и S набраны прописными, так как они обозначают генотипы, а не свойства генов; этого мы будем придерживаться по всей книге). Возвращаясь к начальным данным, можно получить новые частоты генотипов, установившиеся после отбора, из произведения исходных частот и значений W, приведя каждое из них к долям единицы путем деления на сумму этих отдельных членов. Эту сумму иногда обозначают через W, и она равна средней, по всем W.

В табл. 2.3 эти результаты представлены в обобщенном виде.| Из первоначальных частот генов и значений W мы можем теперь! вычислить конечные частоты аллелей (то есть q' и р'). Отсюда час-| тота аллеля а после отбора будет равна
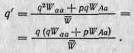
Изменение частоты (А2) за одно поколение равно поэтому q'—q. Приведенные выше уравнения обычно записывают не через W, a через 5, но это лишь обычная замена переменных. Аналогичным образом р' — это просто 1 — q'.
Таблица 2.3. Обобщение результатов вычисления изменений в частотах генотипов, рассмотренных в тексте
| АА | Аа | аа | Всего | |
| Частота до отбора | Р2 | 2рд | 2pq | 1 |
| Приспособленность | Waa | Waa | Waa | |
| Пропорциональный вклад | Waap2 | Waa2pq | Waaq2 | W |
| Частота после отбора в одном поколении | Waap2 W | Waa2pq W | Waaq2 | 1 |
Скорость и характер изменения генных частот от поколения к поколению зависят: 1) от степени доминантности или рецессивности данного аллеля (которая изменяет соотношение значений: W, то есть при доминировании waa= WAa =/= Waa, а в отсутствие доминирования или при сверхдоминировании Waa =/= waa =/= Wqq).
2) от того, какому аллелю благоприятствует отбор (например как уже говорилось, вредный рецессивный аллель может быть скрыт в гетерозиготе, что замедлит его элиминацию; или же благоприятный рецессивный аллель в гетерозиготном состояние будет элиминироваться отбором, что задержит его распространение в популяции); 3) от интенсивности отбора. Использование этих величин приводит к тому, что представленные выше уравнения для изменения частот аллелей приобретают другой вид; вывод всех этих уравнений можно найти в большинстве учебников по популяционной генетике. Однако решения этих уравнений нельзя найти аналитически (то есть исходя из алгебраически уравнений); их можно получить только подстановкой конкретных вещественных чисел вместо q и W (то есть моделированием). Результаты, полученные на одной такой модели, действительно построенной Холдейном, приведены в табл. 2.4. Они относятся к случаю, в котором доминантный аллель обладает преимуществом при относительно низком давлении отбора. Это показывает, что изменение частоты генов зависит от их исходной частоты. При очень низких частотах отбор действует медленнее, чем при промежуточных, а по мере приближения данного аллеля к закреплению (то есть вытеснению другого аллеля) скорость изменения его частоты заметно снижается. Это происходит потому, что менее благоприятный аллель сохраняется в гетерозиготах и его трудно вытеснить полностью. Рецессивному аллелю, обладающему преимуществом, закрепиться при низких частотах еще труднее, поскольку, находясь в гетерозиготном состоянии, он элиминируется из популяции отбором, направленным против менее благоприятного доминантного аллеля, однако закрепление такого рецессивного аллеля происходит быстрее. Ряд моделей, в которых 5> или равно 0,001 и в которых рассматривались разные соотношения между Wгомоз. и Wгетероз., показывает, что сдвиги в генных частотах благоприятных аллелей от 0,1% до более чем 50% особей данной популяции происходят за менее чем 104 поколений. К этому числу мы еще вернемся.
Таблица 2.4. Число поколений, необходимых для того, чтобы произошло да! ное изменение в частоте доминантного аллеля, если доминант оставляют 1000 потомков на 999 потомков рецессивов, т. е Waa = 0,999 или Saa = 0,01
| Р Число поколений | 0,001 до 1% 6920 | 1 до 50% 4819 | 50 до 99% 11664 | 99 до 99,999' 309780 |
Если под действием отбора частота благоприятного аллел возрастает, то отбор должен также повышать среднюю приспособленность популяции в целом (то есть W). Кроме того, поскольку изменение частоты происходит медленнее всего, когда частота одного аллеля очень низка или высока, изменение это, вероятно, будет наибольшим при промежуточных частотах, то есть когда значения частот аллелей наиболее близки друг к другу. Это интуитивная формулировка принципа, который был строго доказан Фишером и считался основной чертой эволюции путем естественного отбора. Его часто называют основной теоремой естественного отбора Фишера, которая формулируется следующим образом: «скорость повышения приспособленности любого организма в любое время равна его генетической дисперсии по приспособленности в это время». Формулировка Фишера неудачна, так как под «организмом» он несомненно имел в виду популяцию. Генетическая дисперсия по W тем больше, чем ближе частоты аллелей и, следовательно, разброс значений W.
Фишер называл свою теорему основной, потому что она строго определяла зависимость между двумя краеугольными камнями дарвиновской теории — изменчивостью внутри популяций и скоростями эволюционного изменения. Однако эта зависимость оказалась не столь несомненной, как полагал Фишер. Так, например, если приспособленность сама зависит от генетической дисперсии (что представляется правдоподобным в некоторых ситуациях; см. разд. 3.5), то скорость изменения приспособленности не должна быть связана с этой дисперсией простой зависимостью. Кроме того, Фишер вывел свою зависимость для случая одного локуса с двумя аллелями; если же рассматривать примеры с участием более чем одного локуса, то зависимость между этими двумя переменными может оказаться значительно сложнее (см. разд. 2.3.3). Есть также и другие потенциальные неясности и проблемы (11). Таким образом, вытекающее из основной теоремы Фишера следствие, что действие естественного отбора всегда направлено на максимизацию приспособленности, нельзя принимать безоговорочно, и из этого в свою очередь вытекает ряд следствий, которые необходимо учитывать при изучении адаптации (см. гл. 3). С основной теоремой Фишера связана еще одна концепция — концепция генетического груза (различие между максимальной потенциальной приспособленностью популяции и W). Это различие создается любым процессом, порождающим генетическую изменчивость, в том числе мутационным процессом (мутационный груз) и процессом генетической перестройки, обусловленной расщеплением (сегрегационный груз). Как и основная теорема Фишера, а возможно, именно из-за нее, концепция генетического груза небесспорна и вызывает разногласия, а поэтому в дальнейшем мы о ней говорить не будем.
Вернемся теперь к вопросам, поставленным в начале этого раздела: насколько отбор важнее, чем повторные мутации? Обратите внимание на то, что для повышения частоты данного аллеля в результате мутирования при отсутствии отбора необходимо, чтобы данная мутация время от времени возникала вновь, то есть повторялась. Рассмотрим мутирование в одном направлении от Л к а, происходящее с частотой и. Вообще обычно происходит также обратное мутирование от а к А, уравновешивающее эффект первого процесса. Однако мы пренебрежем обратными мутациями и используем наиболее благоприятную систему для изменения частоты аллеля путем мутаций. Согласно оценке Холдейна, сделанной на основе наблюдений по дрозофиле и примуле, частота мутаций равна примерно 10~6 на один ген; эта оценка была подтверждена в дальнейшем исследованиями, проводившимися на других организмах. Холдейн вычислил потенциальное воздействие мутаций следующим образом: частота аллеля А в первом поколении потомков (pi) равна (1—и)р<> (где ро — исходная частота Л), а во втором поколении потомков (р2) она равна (1 — и)р! или (1 — и)2р0 и так далее. Записав эти формулы в общем виде, pt=( — и)*р0, и разделив правую и левую части на ро, а затем прологарифмировав, получим
![]()
Теперь мы можем использовать это уравнение для выяснения того, сколько потребуется поколений при частоте мутаций 1:106, чтобы сдвинуть частоту аллеля а (то есть д) с 0,1 до 50%. Если р=—д, то, следовательно, частота аллеля А должна при этом измениться от 99,9 до 50%; поэтому
![]()
Вспомним, что соответствующее значение для действия слабого отбора составляло менее 104 поколений. Таким образом, даже при самых благоприятных допущениях для мутационного процесса и неблагоприятных для естественного отбора последний оказывается гораздо более мощным (почти на два порядка) фактором изменения, чем первый.
Конечно, при этом все еще остается открытым вопрос относительно важности крупных мутаций по сравнению с мелкими. Фишер, как и Дарвин (см. разд. 1.5), утверждал, что вероятность ценности любой мутации обратно пропорциональна ее величине, поскольку вряд ли можно ожидать, что значительное резкое изменение окажется совместимым с другими признаками, с которыми оно должно образовать единое целое. К этому вопросу мы вернемся в гл. 5.
Похожие работы
... матери требовали максимальной "притирки" плода к условиям организма матери, поэтому генетический контроль должен был быть минимальным. 2. Рост мозга. Развитие коры головного мозга. Развитие интеллекта. Эмоциональное развитие. 2.1 Функциональные блоки мозга Дифференциация систем мозговой коры происходит постепенно, и это приводит к неравномерному созреванию отдельных мозговых структур, ...



... . Кроме того, ДНК имеется в митохондриях, которые передаются только по материнской линии. Исследования мутаций в этих ДНК позволяют реконструировать историю биологического развития человечества, происхождение отдельных рас и народностей. Мутация может быть рецессивной, доминантной и полудоминантной в зависимости от состояния гена, в котором она произошла. Гены мутируют с определенной частотой, и ...
— концепция «2К» (кооперация и коммуникация) — эта концепция была разработана в 1900 г. русским ученым-философом П.А.Кропоткиным (1842—1921). В ней идет речь в основном о механизме биологической эволюции. . Термин «эволюция» (лат. evolutio — развертывание) в науке о живом представляет фундаментальное понятие для объяснения возникновения и развития всего живого. Эволюция подразумевает необратимое ...
... о противоречивом пути развития синтетической теории эволюции, о преодолении кризисов в научном познании, направленном на более полное и глубокое понимание закономерностей развития органического мира. Важно отметить, что основы научной теории эволюции заложил Ч. Дарвин. Как господствующее эволюционное учение дарвинизм существовал с 1859 до 1900 гг., т.е. до переоткрытия законов Г. Менделя. До ...
0 комментариев