Навигация
Использование ретровирусных векторов
50401
знак
4
таблицы
0
изображений
1.2 Использование ретровирусных векторов
Результативным способом переноса ДНК в эмбриональные линии животных является применение ретровирусных векторов.
Ретровирусы – семейство эукариотических вирусов, генетический материал которых представлен одноцепочечной РНК.
Вирусы состоят покрытых липопротеидной оболчкой вирусных частиц диаметром 70-120 нм и внутренней капсулы икосаэдрической формы, которая содержит две копии геномной РНК длиной 5-10 тысяч пар нуклеотидов в форме рибонуклеопротеида. Внешняя оболочка вируса является частью цитоплазматической мембраны клетки хозяина и представлена короткими гликопротеидами. Эндогенные ретровирусы являются хромосомными элементами и составляют до 1% ДНК в геноме человека [Tarusio Mantovani, 1998]. Эндогенные ретровирусы являются одной из разновидностей ретроэлементов, составляющих до 10% генома млекопитающих.
По своей способности инфицировать клетки хозяина ретровирусы подразделяются на экотропные и амфотропные. Экотропные вирусы способны реплицироваться в клетках только одного или нескольких близко родственных видов животных. Так, например, вирус лейкемии мыши (MLV) размножается только в клетках мыши и крысы. Амфотропные вирусы, наоборот, имеют широкий спектр хозяев и способны к размножению в клетках различных видов млекопитающих.
Геном всех способных к репликации ретровирусов устроен аналогичным образом и состоит из трех кодирующих регионов, которые, не смотря на то, что речь идет о последовательностях РНК, получили название генов. Ген gag кодирует белки капсида и вирусного кора. Ген pol кодирует вирусную реверсивную транскриптазу и другие связанные с вирусом нуклеазы. Ген env кодирует гликопротеиды в вирусной липидной оболочке.
Геномные РНК ретровирусов содержат на свох концах повторяющиеся последовательности, которые в зависимости от типа вируса имеют длину от 10 до 80 нуклеотидов. Они получили название R-сегментов. На 3' конце геномной РНК R-сегмента расположен регион U3 длиной 170-1250 нуклеотидов, а на 5' конце r-сегмента регион u5 длиной 80-100 нуклеотидов.При образовании ДНК-копии вирусного РНК-генома на концах молекулы происходят изменения: на 5' конце располагается сегмент U3, а на 5' – U5.Такие характерные для ДНК-формы ретровирусоа участки получили название LTR – длинных концевых повторов. LTR, расположенный на 5' конце несет очень сильный промотер, тогда как LTR на 3' конце содержит сигналы полиаденилирования РНК.
Инфекция начинается с взаимодействия ретровируса с клеточной мембраной и связывания поверхностного белка ретровируса (env) со специфическим белком-рецептором. Проникновение ретровируса в клетку происходит посредством микропиноцитоза. У одних вирусов кор, состоящий из белка gag, РНК-зависимой ДНК-полимеразы (реверсивной транскриптазы), интегразы и двух копий ретровирусной РНК, переносится в ядро, где и происходит транскрипция, а у других вирусов транскрипция осуществляется непосредственно в цитоплазме. Обратная транскрипция вирусной РНК в двухцепочечную ДНК осуществляется посредством фермента реверсивной транскриптазы, являющегося частью комплекса ферментов кора. В ходе транскрипции происходит дупликация последовательностей на 5' и 3' концах ретровируса. Если транскрипция имеет место в цитоплазме клетки, то двухцепочечная ДНК вируса транспортируется в ядро, где происходит ее циркуляция с последующей интеграцией одной или нескольких копий в геном клетки. Такая интегрированная ДНК клетки хозяина форма существования вирусного генома получила название провируса. Шаги жизненного цикла ретровирусов.
Для интеграции необходимо наличие на обоих концах ДНК 9 пар оснований, которые узнаются и отщепляются закодированной в вирусе специфической интегразой, являющейся частью комплекса ферментов кора. Процессу интеграции всегда предшествует отщепление двух пар оснований на обоих концах провируса и удвоение 3-6 пар оснований (в зависимости от типа вируса) последовательности ДНК клетки хозяина. Хотя интеграция в геном клетки хозяина не является специфической, следует отметить, что места интеграции часто располагаются в транскрипционно активных участках ДНК. Если происходит интеграция в геном генеративных клеток, то ретровирусы передаются по наследству потомству. В этом случае речь ведут об эндогенных ретровирусах.
Интегрированная вирусная ДНК транскрибируется с помощью РНК-полимеразы клетки-хозяина и трансляцируется как и другие клеточные гены.
Образующиеся в ходе транскрипции продукты являются идентичными вирусной РНК. Все транскрипты содержат на 5 конце кэп-сайт, а на 3' конце участок полиаденилирования. Такие РНК служат материалом для синтеза белков капсида и вирусного кора, а также белков обратной транскриптазы.. Эти белки образуют с геномной РНК вируса комлексы-коры, после чего вирусные частицы покидают клетку через цитоплазматическую мембрану. При этом кор берет с собой часть мембраны клетки хозяина, из которой образует оболочку. Интеграция вирусных частиц в геном клетки хозяина происходит только в митотически активные клетки.
Инфицирование клеток ретровирусами чаше всего не сказывается на их жизнедеятельности. Однако если интеграция провируса произошла вблизи клеточных протоонкогенов, то возможна их активация и, как следствие, гибель клеток. Нарушение жизнеспособности клеток .может иметь место и в том случае интегрирации провируса в действующий клеточный ген (например, опухолевые гены-супрессоры).
Наиболее часто используемыми ретровирусными векторами являюти векторы на основе ретровирусов мыши типа С (вирус лейкемии мыши) Такие векторные системы состоят из двух компонентов: векторнов конструкции и линии клеток-упаковщиц.
В векторной конструкции (провирусная ДНК) гены, кодируюрущие структурные белки ретровируса (gag, pol, env),замешают другими интересующими генами (генами β- галактозидазы и устойчивости к неомицину). Источником структурных белков является клеточная линия, содержащая интегрированный в геном провирус.
Данный провирус модифицируют таким образом, что у него отсутствует сигнал узнавания ψ, необходимый для упаковки вирусной РНК. Таким образом, транскрибируемая вирусная РНК не может быть упакована в вирусные частицы.
РНК векторной конструкции, наоборот, содержит сигнал узнавания ψ и поэтому может быть упакована в рибонуклеопротеидный комплекс. Таким образом, происходит формирование инфекционных вирусных частиц, содержащих информацию ретровирусного вектора и переносящих ее в инфицируемые клетки. Однако такие вирусные частицы не способны к образованию новых вирусов, так как в инфицируемой клеточной линии отсутствует генетическая информация о структурных белках, необходимых для формирования вирусных частиц.
Необходимыми составными частями векторной конструкции являются 5' и 3' LTR, а так же сигнал упаковки, расположенный "вниз по течению" от 5' UTR. Экспрессия трансгена направляется промотором и энхансером 5' UTR или альтернативными вирусными или клеточными промотерами (например, вирус саркомы Рауса, тирозин, α-казеин). Впервые присутствие вирусной ДНК в клетках взрослых мышей установлено в 1974 году. Ретровирусные векторы могут служить альтернативой для эффективного транспорта генов у сельскохозяйственных животных. Инфекция зигот или предимплантационных эмбрионов в большинстве случаев приводит к получению трансгенных животных-мозаиков. Объяснением этому служить то, что интеграция происходит только в том случае, если клетка вступает в митоз после предварительной репликации ДНК. Все потомки, родившиеся из инфицированных ооцитов, были трансгенными {табл. 1). Не смотря на ограниченное число полученных трансгенных животных, были доказаны передача трансгена по наследству и экспрессия рекомбинантного продукта.
Эффективность получения трансгенного КРС с использованием ретровирусной инфекции ооцитов в метафазе II второго мейотического деления.
| Показатели | Число (n) | % |
| Инфицированные ооциты | 836 | |
| Эмбрионы, развившиеся до бластоцисты | 174 | 21 |
| Пересаженные эбрионы | 10 | |
| Живые потомки (% от пересаженных эмбрионов) | 4 | 40 |
| Трансгенные животные (% от родившихся потомков) | 4 | 100 |
Преимуществом использования ретровирусных векторов для получения трансгенных животных является то, что до 100% обработанных эмбрионов могут быть успешно инфицированы ретровирусами.
Недостатком применения ретровирусных векторов является их ограниченная емкость (размер вставки не должен превышать 8 тысяч п. н. Кроме того, в результате сплайсинга из ретровирусов вырезаются интроннные последовательности, которые как и другие дистальные или проксимальные последовательности играют важную роль в эффективной экспрессии генов у трансгенноых животных [Palmiter et ai, 1991]. К недостаткам использования ретровирусных векторов следует так же отнести подавление экспрессии трансгенов in vivo вследствие инактивации вирусных промоторов в клетках, например, посредством а- и у-интерферонов, действующих на вирусные LTR [Ghazizadeh et ai, 1997]. Однако данная проблема может быть решена посредством включения в ретровирусную конструкцию внутренних промоторов или использованием нового поколения ретровирусов, содержащих модифицированные участки контроля экспрессии, такие как внутренний рибосомный сайт (IRES) [Salen, 1997] и тетрациклиновая регуляторная система [lida et ai, 1996].
Еще одним ограничением использования ретровирусных векторов для получения трансгенных животных является их низкий титр. Обычные ретровирусные векторы имеют титр 1 х 105-6 колониеобразующих единиц (cfu) в мл [Kim et ai, 1993; Archer et ai, 1994]. Низкий титр ретровирусов ограничивает способы их введения в эмбрионы. Проблема низкого титра была недавно решена посредством разработки нового вектора на основе вируса везикулярного стоматита (псевдотип VSV-G) [Burns et ai, Yee et ai, 1994]. Известно, что тропизм ретровируса определяется геном env, поэтому его замена в векторе на ген env другого ретровируса может расширить спектр хозяев и изменить свойства ретровируса. Данная технология получила название псевдотипирования. Включение белка вируса везикулярного стоматита С в вектор на основе Мо – MLV (Burns et al,1994) позволило сделать его более стабильным при ультрацентрифугировании. Данный вектор может быть сконцентрирован до титра 1x109-10 КОЕ/мл и имеет очень широкий спектор клеток-хозяев. С его использованием были получены трансгеные лини клеток насекомых, млекопитающих, амфибий, рыб, а также созданы трансгенные животные. Высокий титр ретровируса сделал возможным получение трансгенных животных посредством инъекции содержащей вирусы среды в перивителиновое пространство [Chan et al, 1998)/ В ооцитах крупного рогатого скота объем перивителинового пространства составляет 200-300 пкл. При использовании вируса с титром 1*10 9-10 КОЕ/мл, инъекция 10 пкл приводит к проникновению в перивителиальное пространство от 10 до 100 инфекционных вирусных частиц. Если титр вируса равен 1*105-6 КОЕ/мл, то для введения в эмбрион одной вирусной частицы понадобилось не менее 1 нл раствора .
К недостаткам использования ретровирусов относится возможность клеточных онкогенов посредством вирусных транскрипционных последовательностей. Даже при применении ретровирусов, которые не могут существовать как инфекционные вирусные частицы, существует хотя и очень небольшой, но риск, что при рекомбинации с присутствующими в эмбрионах эндогенными ретровирусными последовательностями могут образовываться новые активные формы ретровирусов.
Не смотря на перечисленные недостатки и ограничения, ретровирусы могут быть широко использованы для трансгенеза в животноводстве. Успешное введение генетического материала в ооциты делает возможным проведение генной терапии наследственных заболеваний. Использование ретровирусных векторов позволяет осуществлять трансформацию отдельных органов. Так, Archer с соавторами [1994] ввели в молочную железу козы путем инфузии через сосковый канал в период гормонально индуцируемого маммогенеза ретровирус, кодирующий гормон роста человека (hGH). Лактация наступила на 14-ый день гормональной обработки. Максимальный уровень hGH (60 нг/мл) наблюдался в первый день лактации и затем опускался до 12 нг/мл на 9-12 дни лактации.
Аналогичные эксперименты с использованием экспрессионного вектора pLNS, полученного на основе Mo-MLV проводятся в отделе биотехнологии Всероссийского НИИ животноводства.
LTR – длинный концевой повтор , ψ+ - сигнал упаковки, Н4 – промотер гистона человека, NEO – ген устойчивости к неомицину,ЕР – делеция в области промотера-энхансера, рА – сигнал полиаденилирования Данный вектор содержит только цис-действующие последовательности генома вируса, необходимые для репликации. Последовательности вируса, кодирующие белки, исключены из векторной конструкции. Вектор содержит 5' длинный концевой повтор LTR, с которого происходит транскрпция, ψ+ область, ответственную за упаковку вирусного генома в вирион, ген устойчивости к неомицину, под контролем промотера Н4 гистона человека для селективного введения конструкции в клеточную линию и 3' LTR с делецией в области промотера-энхансера вируса. После одного цикла репликации делеция в области ЕР LTR будет присутствовать на обоих концах провируса, регуляторные элементы провируса будут полностью инактивированы и клонированные гены будут экспрессироваться только под контролем собственных промотеров.
Доказана возможность успешной инфекции альвеолярных клеток молочной железы свиней и коров рекомбинантными ретровирусами..
Инфекция молочной железы не носит локального характера. Последовательности провируса помимо опытных долей вымени были выявлены в контрольных долях молочной железы, в слюнной железе, селезенке, поджелудочной железе, спинном мозге, надпочечниках. ИФА молока свиней выявил наличие рекобинантного продукта в количестве от 30 до 290 нг/мл.
Основными недостатками метода непосредственной инфекции отдельных органов животных является необходимость использования большой дозы ретровирусов с тем, чтобы инфицировать достаточное число клеток, а так же невозможность передачи трансгена по наследству следующему поколению животных. Однако в отличие от введения грансгенов в эмбриональные линии, данный подход позволит синтезировать рекомбинантные продукты в молоко уже через несколько месяцев после начала эксперимента. Для КРС по сравнению с традиционным методом микроинъекции затраты времени от начала эксперимента и до получения первых результатов об уровне экспрессии в молоке сокращаются, соответственно, с 4-5 лет до 4 5 месяцев.
Похожие работы
... например, "людей-свиней". Но есть и более логичные доводы. Скажем, новые медицинские и биологические препараты разрешаются к использованию на людях только после многолетних проверок на животных. Трансгенные продукты поступают в свободную продажу и уже охватывают несколько сотен наименований, хотя созданы они были всего несколько лет назад. Противники трансгенов подвергают сомнению и методы оценки ...
... , вызванные динамическими му-тациями.-----------------------T-----------T-------T-----T------T------T----------------------¬ Болезнь, номер по ¦ Ген, лока-¦Триплет¦Норма¦Прему-¦Мута- ¦Литература ¦ МакКьюсику (MIM) ¦ лизация ¦ ¦ ¦тация ¦ция ¦ ¦ -----------------------+-----------+-------+-----+------+------+----------------------+ Синдром ломкой X-хро- ¦FMR1, FRAXA¦(CGG)n ...

... при постановке биологических или медицинских экспериментов. Составлять его должны ведущие специалисты в этой области и знающие предмет юристы. 3. Другие применения генной инженерии Несмотря на успехи ученых в генной инженерии в таких отраслях, как создание генно-модифицированных продуктов и клонировании животных и человека, они на этом не останавливаются. В целях сохранения военного ...
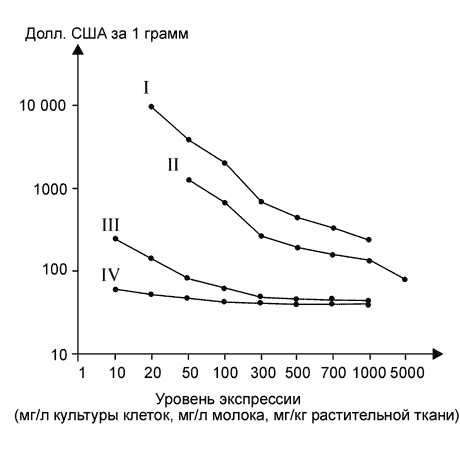
... растений позволит значительно снизить её стоимость (Giddings et al., 2000). В заключение хотелось бы отметить, что несмотря на значительные достижения в области продукции реком-бинантных белков медицинского назначения в растениях, это направление находится лишь на начальном этапе своего развития. Учёные-биотехно-логи уверены, что в будущем рекомбинантные препараты, получаемые из генетически ...
0 комментариев