Основные болезни и вредители сои
Регуляторы роста и развития растений, как элемент технологии повышающий устойчивость растений к стрессам
УСЛОВИЯ И МЕТОДИКА ПРОВЕДЕНИЯ ИССЛЕДОВАНИЙ
Почвенно-климатические условия
Материалы и методы исследований
Влияние регуляторов роста растений на фитосанитарное состояние посевов сои сорта Вилана: учеты проводились по общепринятым методикам
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ И ИХ АНАЛИЗ
Влияние обработок регуляторами роста растений на фитосанитарное состояние сои сорта Вилана
Г, сорта Вилана болезнями, фаза цветения, ВНИИМК
Влияние обработок регуляторами роста растений на урожай и
БЕЗОПАСНОСТЬ ЖИЗНЕДЕЯТЕЛЬНОСТИ
ОХРАНА ОКРУЖАЮЩЕЙ СРЕДЫ
Пропаганда природоохранительных знаний с увязкой задач сельскохозяйственного производства и охраны природы в целом [6]
Навигация
Регуляторы роста и развития растений, как элемент технологии повышающий устойчивость растений к стрессам
Влияние регуляторов роста растений на физиолого-биохимические показатели урожайности сои
141752
знака
25
таблиц
25
изображений
1.3. Регуляторы роста и развития растений, как элемент технологии повышающий устойчивость растений к стрессам
Одним из факторов ведущим к загрязнению окружающей среды является применение необоснованно, большого количества средств защиты растений. В связи с этим актуальным является применение интегрированной защиты растений, которая подразумевает получение запланированного урожая высокого качества. Для этого необходимо оптимальное сочетание факторов урожайности: плодородие почвы, питание растений, качественная сельскохозяйственная техника и технология возделывания, место произрастания, селекция устойчивых сортов и гибридов, севооборот, оперативная защита растений.
Одним из агроприемов, повышающих устойчивость растений к стрессам, а так же урожай и его качество у многих сельскохозяйственных культур, может считаться применение стимуляторов роста растений.
Основные классы фитогормонов: в роле стимуляторов роста растений выступают фитогормоны – это вещества, вырабатывающиеся в процессе естественного обмена веществ и оказывающие в ничтожных количествах регуляторное влияние, координирующее физиологические процессы. В этой связи к ним часто применяют термин – природные регуляторы роста. Гормоны способны к передвижению по растению и их влияние носит дистанционный характер.
Большинство физиологических процессов, в первую очередь рост, формообразование и развитие растений, регулируется гормонами. Гормоны играют ведущую роль в адаптации растений к условиям среды. К фитогормонам, стимулирующим рост и развитие растений, относят: ауксины, гиббереллины, цитокинины. По мнению Аноп (1986) действие регуляторов роста растений кратко можно охарактеризовать по группам:
Ауксины – обуславливают быстрое деление клеток растений, но при завышенных нормах могут привести растение к гибели;
Гиббереллины – вызывают существенное изменение отдельных признаков растений, например: длины стебля, всхожести семян, сроков цветения и т.д.;
Этилен – широко применяют для ускорения созревания плодов, а также для нарушения покоя у картофеля;
Ксантоксин – у пшеницы и зернобобовых способствует снижению транспирации по средствам регулирования закрытия устьиц (в настоящее время регуляторы этого типа широко используют для десикации хлопчатника;
Цитокинины – стимулируют синтез белков у растений, а так же увеличивают продолжительность фотосинтетической деятельности листьев.
Ауксины – это вещества индольной природы. Основным фитогормоном типа ауксина является β-индолилуксусная кислота (ИУК).
Учеными: Ч. Дарвиним, П. Бойсен – Йенсеным, Вентом в разное время были проведены исследования и опыты, которые показали, что в верхушке проростков вырабатывается особое вещество, которое, передвигается к нижележащим клеткам, регулирует их рост в фазе растяжения. И поскольку это вещество вырабатывается в одной части растения, а вызывает физиологические эффекты в другой, оно было отнесено к гормонам роста растений – фитогормонам. Исследования показали, что рост различных видов растений, и отдельных органов регулируется ауксином – это широко распространенный стимулятор роста и развития растения. Наиболее богаты ауксинами растущие части растительного организма: верхушки стеблей, молодые растущие части листьев, почки, завязи, семена и пыльца. Образование и наличие ИУК зависит от снабжения растения азотом, обеспечения растений водой. Освещение уменьшает содержание ауксинов, а затемнение увеличивает. Под влиянием микроорганизмов содержание ауксинов у высшего растения заметно возрастает. Через изменение содержания фитогормонов осуществляется первоначальное влияние условий внешней среды на процессы обмена веществ и рост. Содержание ауксинов меняется в процессе онтогенеза растения. Регуляция образования и разрушения ИУК – это один из способов регуляции ее содержания, а, следовательно, и процессов роста.
Физиологические проявления действия ауксинов: наиболее ярким проявлением физиологического действия ауксина является его влияние на рост клеток в фазе растяжения. Под действием ауксинов индуцируется корнеобразование, вызывается дифференциация ксилемы. Большую роль ауксины играют при разрастании завязи и плодообразовании. Действие ауксина находится в зависимости от его концентрации. Повышение концентрации ауксина выше оптимальной вызывает торможение роста. Одна и та же концентрация его может усилить рост одних органов и затормозить рост у других. При всех физиологических проявлениях ауксины усиливают поступление и перераспределение воды и питательных веществ в растение. В 1933 году появились исследования, показывающие, что под влиянием ауксина интенсивность дыхания растет. ИУК увеличивает энергетическую эффективность дыхания растений. Под влиянием ИУК возрастает и энергетический заряд клетки (отношение АТФ + АДФ к АМФ).
Гиббереллины – широко распространенные среди растений вещества, обладающие высокой физиологической активностью и являющиеся фитогормонами. В настоящее время известно около 70 веществ, относящихся к группе гиббереллины: ГА1; ГА2 и другие. Растения на разных этапах онтогенеза могут различаться по набору гиббереллинов, активность которых может быть различной. Основное место образования гиббереллинов – это листья.
Гиббереллины существуют в двух формах: свободной и связанной. Они могут передвигаться как вниз по растению, так и вверх. Освещение увеличивает содержание гиббереллинов, улучшение питания растения азотом уменьшает содержание гиббереллинов в растение, уменьшение влажности почвы снижает содержание гиббереллинов. Большое содержание этого фитогормона находится в период прорастания семян. Содержание его в процессе онтогенеза в листьях изменяется в соответствие с одновершинной кривой, возрастая вплоть до цветения, а затем уменьшаясь. Некоторые исследователи считают гиббереллин гормоном роста: он резко усиливает рост стебля у карликовых форм различных растений, увеличивая длину стебля на 30-50 %. Увеличение роста осуществляется как за счёт деления клеток, так и за счёт их растяжения. Гиббереллины участвуют в разрастании завязи и образовании плодов. Они накапливаются в почках при выходе из покоящегося состояния. В соответствие с этим обработка гиббереллином вызывает прерывание покоя у почек. Гиббереллины способствуют накоплению питательных веществ, возрастает общая масса растительного организма. Гиббереллины накапливаются в хлоропластах и оказывают большое влияние на процессы фотосинтеза, усиливая процессы фотосинтетического фосфорелирования, при этом снижается содержание хлорофилла и повышается интенсивность использования единицы хлорофилла, возрастает ассимиляционное число. В темноте гиббереллин воздействует лишь на растяжение клеток, не вызывая возрастания интенсивности их деления.
В 1955 г. было выделено активное начало, вызывающее деление клеток: 6-фурфуриламинопурин, в растениях это вещество не встречается, но были найдены близкие химические соединения, регулирующие процесс деления клеток – цитокинины. Все известные цитокинины – это производные пуриновых азотистых оснований, а именно аденина, в котором аминогруппа в шестом положении замещена различными радикалами. Соединения цитокининового типа в растениях находятся не только в свободном состоянии, но и в составе некоторых т-РНК. Цитокинины образуются в основном в корнях и передвигаются в наземные органы по ксилеме. Цитокинины во многом определяют физиологическое влияние корневой системы на обмен веществ надземных органов. Цитокинины могут развиваться так же в семенах и плодах.
Улучшение питания растений азотом усиливает образование цитокининов. Вместе с тем для проявления их действия необходимо достаточное снабжение растений питательными веществами, особенно азотом. Цитокинины активируют рост растяжением, так же они способствуют пробуждению и росту боковых почек.
Цитокинины оказывают влияние на ультраструктуру хлоропластов, они повышают содержание хлорофилла, ускоряя образование его предшественника – протохлорофиллида, так же они усиливают интенсивность фотофосфорелирования, вместе с тем активируют синтез ключевого фермента темновых реакций фотосинтеза – РДФ – карбоксилазы. Цитокинины усиливают передвижение веществ к обогащенным ими тканям (Якушкина, 1993).
Регуляторами роста растений называются физиологически активные вещества биологического происхождения или синтезированные искусственно, воздействующие на интенсивность и направленность процессов жизнедеятельности растений, позволяющие им более эффективно использовать все, что запланировано генотипом растений, но в силу ряда причин осталось нереализованным. Регуляторы роста дают возможность повышать их урожай, улучшать качество, условия уборки и хранения продукции (Казакова, 1986)
В настоящее время обнаружено и в разной степени изучено более 4 тысяч биологически активных веществ, 10 % из которых нашли применение в сельскохозяйственном производстве. Наиболее эффективными регуляторами роста, применяемыми в сельском хозяйстве, являются: кампозан М, гиббереллин. В России внедряются всего 10 препаратов на 24 культурах для обработки семян и опрыскивания растений, с целью предотвращения полегания и повышения урожайности. Большинство применяемых ныне регуляторов роста, как правило, влияют на укорачивание стебля, предотвращение предуборочного опадения плодов, ускорение созревания, и облегчают уборку урожая (Oplinger, 1985).
Условно действие регуляторов роста на физиологические процессы в растениях можно разделить на два типа: индукцию и стимуляцию. Индукция представляет собой включение под действием регуляторов роста процесса, который не происходит в клетке в отсутствие данного фитогормона, тогда как стимуляция сводится к усилению, активации уже идущих в клетке процессов. Такое разграничение необходимо проводить при изучение механизма действия регуляторов роста, так как молекулярные механизмы индукции и стимуляции различные. Вместе с тем в жизни клетки эти два механизма могут, по – видимому проходить одновременно и в ряде случаев трудно провести четкую грань между их проявлениями (Кулаева, 1977).
Влияние экзогенных обработок регуляторами роста растений на рост и развитие сельскохозяйственных растений: урожайность зависит от избирательной способности растения направлять энергию, обычно расходуемую на вегетативный рост, на формирование семян. Эффективность этого процесса – один из главных факторов определяющих урожайность. Это особенно наглядно для растений сои, которые формируют намного больше вегетативной массы, чем это необходимо для формирования семян. Регулятор роста, подавляющий излишний вегетативный рост, позволил бы направить больше энергии на репродуктивные процессы и образование большего числа цветков, создавая при этом потенциал для повышения продуктивности (Никелл, 1984)
Howell K.W. (1963) при опрыскивание растений сои в полевых условиях гиббереллиновой кислотой (ГК) уже на второй день наблюдал увеличение высоты растений, а при предпосевной обработке семян сои ГК отмечал более быстрое появление всходов. Несмотря на полученные результаты, влияние предпосевной обработки семян и опрыскивание листовой поверхности не привело к увеличению урожайности. Автор заключает, что применение ГК на посевах сои экономически не оправдано. Однако, в опытах проведенных Бугаковой А.Н. (1986) отмечено, что предпосевное намачивание семян сои ГК привело к стимулированию ростовых процессов от появления всходов до созревания. Кроме того, обработка семян ГК способствовала увеличению числа и массы бобов.
Bostrack J. M., Struckmtyer B. E. при опрыскивании сои раствором ГК в концентрации 50 мг/л не отметили существенных различий в форме и размере прилистников, но отмечали уменьшение размеров листовых пластинок и на них хлоротичные пятна. Удлинение клеток вызывало преждевременное удлинение междоузлий у обработанных растений. Уменьшение диаметра клеток при этом приводит к уменьшению диаметра стебля. Удлинение боковых ветвей у обработанных ГК3 растений несколько задерживалось, но затем они превосходили по росту контрольные. У обработанных ГК3 растений в условиях длинного дня наблюдали уменьшение количества бобов. При этом нужно отметить, что у таких растений переход от вегетативной к репродуктивной фазе затягивается. Однако, в опытах Бугаковой А.Н. (1986) предпосевное намачивание семян сои ГК3 (100 мг/л) привело к ускорению развития, и растение зацвело на неделю раньше, чем в контроле, и на 4-6 дней раньше, чем в варианте с ГК3 (10 мл/л).
Ounsworth L. E., Pillay D. T. M. (1969) на фоне различных доз удобрений испытывали влияние ГК3 и Диметил гидразид янтарной кислоты (ДГЯК) на ростовые процессы и поглощение NPK растениями сои. Авторы отмечали, что обработка растений ГК3привела к увеличению высоты растений, тогда как действие ДГЯК оказалось противоположным. К аналогичным выводам при испытании ДГЯК пришел и Gowda P. M. (1972)
Bouniols A. и другие, (1978) при возделывании сои сорта «Amsoy 71» с различными густотами стояния (от 50 до 800 тыс. растений/га) на орошение и без него изучали влияние опрыскивания растений различными дозами ДГЯК и ССС в фазу четырех листьев и на пять дней позже. Оба регулятора роста уменьшали высоту растений на более загущенных посевах (более 300 тыс. растений/га), увеличивая при этом количество цветков и бобов вследствие снижения процесса опадения репродуктивных органов. Обработка растений хлорхолин хлоридом улучшило качество белка в семенах. Наибольший эффект отмечен в богарных условиях.
Эндогенные гормоны играют важную роль в регулирование опадения цветков и бобов. Общеизвестно, что этилен, абсцизовая кислота и некоторые ауксины способствуют более быстрому протеканию этого процесса, тогда как гиббереллины, цитокинины и другие типы ауксинов значительно уменьшают его; влияние различных типов ауксинов зависит от их способности индуцировать синтез этилена (Porter, 1977, Nooden, Leopold, 1978, Sagee et al., 1980, Wagenbreth et al., 1981, Sexton, Roberts, 1982)
Опрыскивание сои гибберелловой кислотой оказало положительное влияние на формирование бобов у сои сорта Вильямс в условиях Кубы, однако оно привело к уменьшению сухой массы семян, но формирование большого количества бобов положительно сказалось на продуктивности растений. При этом опадение цветков и бобов у контрольных растений достигло 66,3, тогда как при опрыскивание гибберелловой кислотой в дозе 5 и 50 мкг/растение опадение составило соответственно 52,8 и 48,1 % (Castro, 1985).
Согласно Wiebold (1981) наибольшее опадение бобов происходит до того, как они достигают 2 см длины.
При изучение процесса опадения цветков и бобов у трех сортов сои обработанных Г3К, Castro (1985) пришел к выводу, что этот процесс оказался идентичным по всем сортам и, что раннее опадение бобов происходит из-за низкого содержания эндогенных гиббереллинов и цитокининов.
Meade J. A. (1958) установил, что обработка растений сои изопропил N (3–хлор фенил) карбоматом вызывает увеличение содержания редуцирующих сахаров в листьях до 35 % с одновременным их уменьшением в стеблях до 60 % от обычного их содержания. Инвертирующие сахара в листьях обработанных растений было в 14 раз выше, чем у контрольных растений. Общее содержание сахаров у обработанных растений оказалось на 90 % больше, чем у контрольных растений.
Oizume H. (1962) установил, что обработка растений в начале их роста гидрозид малейновой кислотой (ГМК) приводила к остановке роста стебля. Формирование боковых значительно увеличивалось. При этом приходит к выводу, что появление боковых побегов в основном зависит от содержания доступных углеводов в растениях и, что содержание азота не является лимитирующим фактором их появления.
Hew et al. (1967) установил, что при опрыскивание растений сои ИУК и Г3К наблюдается увеличение в транспорте сахарозы.
Cothren J.J., Stutte C.A. (1973) при изучение влияния опрыскивания сои в контролируемых условиях хлор-3-бутил фосфоником и тетрагидро-фурфурил изотиоцианатом в концентрациях 250 мг/г на содержание сахаров в листьях сои установили, что регуляторы роста снижают процентное содержание глюкозы и фруктозы и увеличивали содержание сахарозы. В полевых условиях обработка растений тетрогидро-фурфурил изотиоцианатом привело к увеличению содержания общего сахара в листьях сорта сои Davis, тогда как содержание глюкозы, фруктозы и сахарозы находилось на одном и том же уровне что и при выращивание сои в контролируемых условиях.
Определение осмотического потенциала сельскохозяйственных культур, предоставляет возможность узнавать наиболее оптимальные гидрологические условия для роста и развития. Морфологические и физиологические изменения, вызываемые регуляторами роста растений, могут привести к положительным изменениям гидрологического равновесия растения и таким образом к увеличению урожая. Так в опытах Cothren J.T., Rutledge S.R., Stutte C.A. (1975) установлено, что применение регулятора роста в условиях орошения сорта сои R 56-49, Davis, Hale-7 привело к увеличению урожайности от 16 до 38 %, тогда как в богарных условиях повышение урожайности составило всего 7-23 %.
Таким образом, важность типа растений (детерминантного или индетерминантного) проявляется в том, что их физиологические реакции находятся под контролем внешней среды. Химические вещества могут быть эффективны на детерминантных типах сои, и не влиять на индетерминантные типы, и наоборот.
Stutte C.A. (1973, 1974, 1975, 1976) разработал обширную программу скрининга и оценки регуляторов роста растений по их потенциальному влиянию на сою. Результаты этих испытаний показали положительное влияние следующих препаратов: хлористый трибутил (5-хлор-2-тиэнил) фосфоний, тетрогидрофурфурил изотиоционат и 1,2-дигидро-4,6-диметил-2-оксоникатиновая кислота.
При обработке семян гороха БИРР за 10 дней до посева показала преимущество дозы 2 мл/т, линейные проростки увеличились на 2-27,9 %. При первом замере вегетирующих растений гороха, обработанные семена были выше не обработанных на 31,4-34,1 %, но в последствие они выровнялись и стали вегетировать примерно на одном уровне. Следовательно, БИРР увеличивает рост растений на первоначальных стадиях развития растения (Офицерова, 2006) .
Влияние экзогенных обработок регуляторами роста растений на устойчивость сельскохозяйственных культур к стресс факторам. Обработанные растения становятся более устойчивыми к неблагоприятным условиям среды, способствуют выходу из покоящегося состояния спящих почек, семян, клубней (Якушкина, 1993).
Изменение условий окружающей среды (температура, влажность, освещение и минеральное питание) вызывают изменения в активности всех фитогормонов. При этом часто активность отдельных гормонов изменяется в противоположных направлениях.
Влияние неблагоприятных условий среды можно смягчить путем экзогенного введения гормонов. Показательными в этом отношении являются данные по влиянию обработки гиббереллином проростков, выращиваемых в условиях повышенной температуры. Внесение ГК3 (гиббереллиновая кислота) восполняет дефицит эндогенных фитогормонов и этим самым устраняет подавляющее влияние повышенной температуры на рост растения.
Альбит повышает эффективность применения гербицидов. Гербициды на 50 % могут снижать урожайность, поэтому применяют их с препаратами антистрессантами. Альбит снижает стресс от гербицидов на 5-38 %, снижает восприимчивость обработанных растений к болезням (мучнистой росе, бурой ржавчине, пятнистостям). За счет своей ростостимулирующей активности и снятия стрессового эффекта гербицида Альбит повышает урожайность на 4,7 ц/га, повышает качество урожая (клейковина у зерновых на 0,5-5,1 %, сахаристость у сахарной свеклы, содержание витаминов в овощах), защищает растения от основных болезней (средняя биологическая эффективность – 40-80 %); позволяет снизить расход протравителей и фунгицидов на 25-100 %, удобрений на 30-40 %; снимает гербицидный стресс у основной культуры, что повышает устойчивость растений к листостеблевым инфекциям и позволяет сохранить до половины урожая; усиливает засухоустойчивость растений на 10-60 %. (Злотников, 2006)
Эффективность обработки семян стимулятором роста БИРР. Обработка семян биологически активными препаратами и опрыскивание ими посевов позволяет повысить биологическую активность семян, активизировать физиологические процессы во время вегетации растений, повысить их адаптационные возможности в неблагоприятных условиях и продуктивность, улучшить качество выращиваемой продукции. Масса корней при обработке БИРР была больше на 33,3 %. Обработка БИРР позволила снизить заболеваемость растений корневыми гнилями. Не обработанные растения заболели на 100 % со степенью развития 45 %, а обработанные поразились на 79 %. Полевая всхожесть повысилась на 9 %, урожайность на 13,2 %. Обработку следует проводить в дозе 2 мл/т в срок от одного дна до 20 дней до посева (Офицерова, 2006).
Применение регулятора роста Силк на картофеле: воздействуя на обмен веществ, регуляторы роста способствуют росту и развитию растений, стимулированию иммунитета, устойчивости ко многим болезням грибного, бактериального и вирусного происхождения. Результат их взаимодействия – увеличение продуктивности и хозяйственно-биологической оценки полевых культур. Силк испытывался как клубневой и листовой регулятор. Тестовый анализ показал преимущество биорегулятора Силк на картофеле в улучшение урожайных (прибавка 3,2-9,2 т/га или 14,5-37,1 % в зависимости от способа применения), товарных (рост коэффициента размножения на 1,2-3,4; товарности на 20-24 % при опрыскивание листьев) и технологических (увеличение крахмала на 1-3,6 %; витамина С на 10-14 мг) свойств.
При замачивание клубней в растворе Силка повышается вегетативная масса: высота стеблей на 15 см, число стеблей в 2-3 раза, площадь листьев и сырой массы ботвы в 2 раза. Это приводит к снижению чистой продуктивности фотосинтеза. Наибольший эффект имела одинарная норма (10 мл действующего вещества/т + 10 мл действующего вещества/га). (Засорина, 2006).
В последнее время в качестве иммунизаторов используют регуляторы роста растений. Так, парааминобензойная кислота при обработке вегетирующих растений, повышает их устойчивость к заболеваниям и стресс факторам. Обработка вегетирующих растений пшеницы иммунизатором и регуляторами роста растений – фуроланом повышает урожай зерна и снижает поражение растений фузариозом колоса, корневыми гнилями, септориозом, бурой ржавчиной. Индукторы устойчивости мивал, крезацин, эль, иммуноцитофит, хитозан, силк, амбиол, агат-25, эмистим С активируют естественные защитные механизмы растений, снижают поражение корневыми гнилями, бурой ржавчиной, при этом крезацин и гуминовые кислоты существенно повышают урожай зерна (Андрианова, 1999).
Эпин повышает устойчивость растений огурца к пероноспорозу, особенно у малоустойчивого сорта в условиях эпифитотии пероноспороза, увеличивает интенсивность окислительных процессов при развитии заболевания (Балахонцев, Исхаков, 1999; Заплатин, Хрянин,1999).
Похожие работы
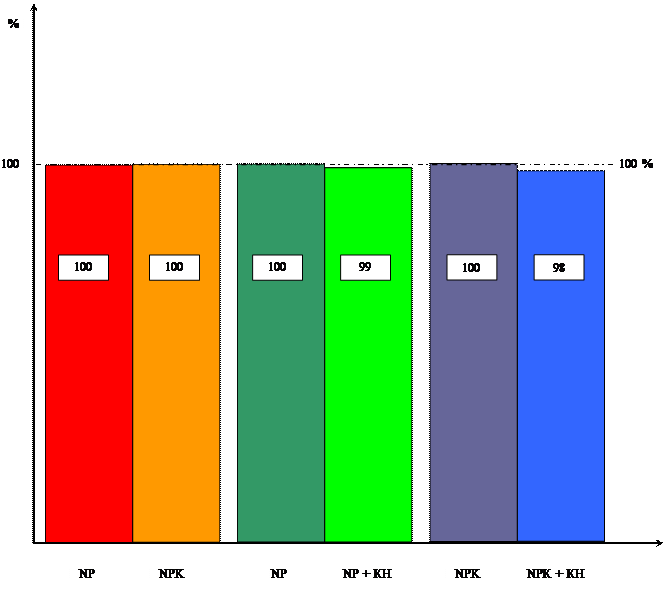
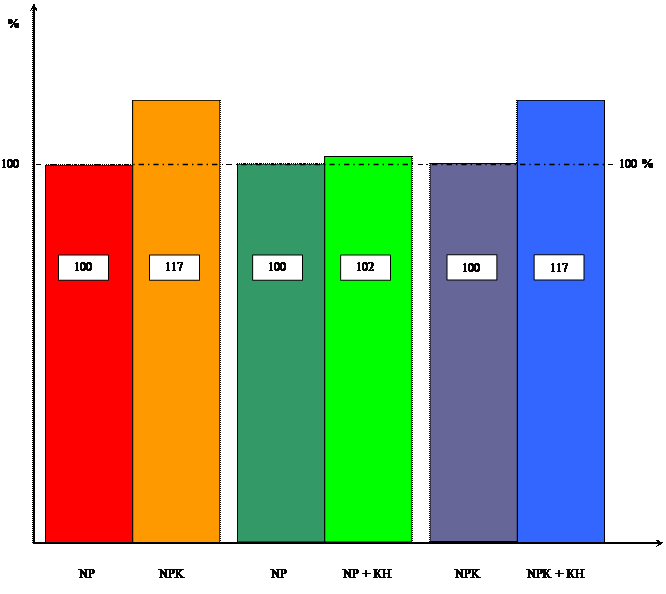
... системы, но вызвал увеличение ее массы, мы провели исследования по влиянию кинетина на накопление массы корневой и побеговой систем в зависимости от уровня минерального питания. Данные расположены в таблицах 3 и 4. Таблица 3 Влияние кинетина на массу побеговой системы 10-ти проростков пшеницы в зависимости от уровня калийного питания ( мг). Возраст, дн. NP NP + КН % NPK NPK + КН % ...

... фазы развития. Кроме того, применение указанных гаметоцидов задерживало прохождение фенологических фаз у растений, а также выколашивание части стеблей. Однако, несмотря на отрицательные побочные действия гаметоцидов, применение их в ближайшей перспективе может открыть путь к реализации эффекта гетерозиса у зерновых (особенно пшеницы), технических, овощных и кормовых культур. Поэтому весьма ...
... . У засухоустойчивых растений эти показатели будут выше. В селекционной работе используют такой показатель, как содержание статолитного крахмала в корневом чехлике. Генетически обусловленным признаком засухоустойчивости растений является способность их вегетативных органов (особенно листьев) накапливать во время засухи пролин. При этом концентрация пролина увеличивается в 10—100 раз. В пролине ...
... к селекционерам, как непременная устойчивость сортов к механическим повреждениям, повергает последних в глубокое уныние (а подобных требований к перерабатываемому сорту десятки) . Вот почему биотехнологи стремятся как-то помочь старым апробированным сортам — улучшить их продуктивность, качество или товарный вид. Привередливые американцы, например, предпочитают апельсины ярко-оранжевой окраски. ...
0 комментариев