МОСКОВСКИЙ ПСИХОЛОГО-СОЦИАЛЬНЫЙ ИНСТИТУТ
РЕФЕРАТ ПО ПСИХОФИЗИОЛОГИИ НА ТЕМУ:
Гамкергическая теория депрессии у человека
Студентка:
КондратьеваД.А.
Группа:
22ПМ/4-08
Преподаватель:
МОСКВА 2006
Содержание
Биологические закономерности депрессии………………….………….3
ГАМК, тревога и депрессия………………………………………………5
Заключение……………………………………………………………….11
Литература………………………………………………………………..14
Биологические закономерности депрессии.
В последние годы актуальными являются исследования, связанные со стрессом и определением его вклада в патогенез невротических и аффективных расстройств.
Выявлено, что при депрессивных расстройствах наблюдаются супрессия Т-клеточного иммунитета, снижение количества натуральных киллеров, дисрегуляция факторов неспецифической резистентности, активация гуморальных факторов иммунитета. Иммунные нарушения сопряжены с изменением нейроэндокринной регуляции, проявляющейся повышением уровня кортизола и активацией нейромедиаторного обмена. Различный характер иммунобиологического реагирования соотносится с особенностями клинического полиморфизма аффективных расстройств. Биполярные аффективные расстройства характеризуются наиболее значимыми нарушениями. В то же время, характерной особенностью психонейроиммунных взаимодействий при хронических аффективных расстройствах (дистимии) является высокая активность параметров нейромедиаторного обмена (серотонина и дофамина) при нормальной концентрации гормонов. Использование кластерного анализа позволило выявить ученым взаимосвязи клинических характеристик депрессивных расстройств с иммунологическими, нейромедиаторными и гормональными показателями, что доказывает вовлеченность этих гомеостатических систем в механизмы психонейроиммуномодуляции и в патогенез депрессивных расстройств. Степень выраженности депрессивной симптоматики сопряжена с параметрами нейромедиаторного обмена, концентрацией адренокортикотропного гормона, количеством хелперов/индукторов, а длительность заболевания коррелирует с количеством цитолитических киллеров/супрессоров, натуральных киллеров, В-лимфоцитов.
Таким образом, основные закономерности психонейроиммуномодуляции при депрессии проявляются дисбалансом иммунной, нейромедиаторной и гормональной систем.
Рассматривая общие нейрохимические, фармакологические и нейрофизиологические аспекты тревоги и депрессии, ученые формулируют концепцию о взаимосвязи тревоги и депрессии в единый патогенный процесс.
Стресс является ведущим фактором патогенеза ЦНС, в первую очередь тревоги и депрессии - наиболее распространенных заболеваний психики [35, 50,51,52]. Гетерогенность механизмов патогенеза и форм тревоги и депрессии, высокий процент коморбидности, а также сходство многих клинических проявлений существенно затрудняет диагностику и терапию данных патологий [22,52]. За последние десятилетия выдвинуто множество гипотез о механизмах возникновения тревоги и депрессии [35,72], однако полная картина их патогенеза еще не известна. Наличие одной из указанных патологий осложняет терапию другой патологии, является предиктором плохого терапевтического результата и фактором риска возникновения других заболеваний ЦНС [50,51]. Это, а также сходство клинических симптомов тревоги и депрессии, их генетических особенностей и общая зависимость патогенеза от стресса указывают на то, что данные патологии взаимодействуют друг с другом, а при возникновении тревоги и депрессии могут быть задействованы общие механизмы, биологические маркеры и факторы риска [52].
Анализ клинической эффективности ряда антидепрессантов (АД) и анксиолитиков (АЛ) также подтвержает взаимосвязь тревоги и депрессии, поскольку синергический терапевтический эффект при монотерапии оказывают многие препараты обоих классов, а сочетание АД и АЛ часто оказывает эффект, превосходящий сумму эффектов каждого из них в отдельности [52]. На возможную общность генеза тревоги и депресии указывают данные из области эволюционной психиатрии [47]. Немаловажным аргументом в пользу общности тревожно-депрессивного патогенеза служат обширные сведения из области экспериментальной фармакологии тревоги и депрессии, которые будут рассмотрены ниже. Таким образом, в современной биологической психиатрии необходимы новые подходы к пониманию общих механизмов возникновения тревоги и депрессии.
Гамма-аминомасляная кислота (ГАМК) – важнейший тормозный медиатор мозга, задействованный во многих процессах ЦНС, включая регуляцию эмоций, поведения, памяти и высших когнитивных функций [35,46]. Существующие клинические и экспериментальные данные позволяют допустить, что именно ГАМК может быть тем связующим звеном, нарушение которого приводит к патогенезу как тревоги, так и депрессии, их коморбидности, а также объясняет сходство симптомов и чувствительность к одним и тем же препаратам.
ГАМК, тревога и депрессия.
В течение десятилетий в психофармакологии доминировали аминергические теории патогенеза депрессии, а патогенез тревоги традиционно объяснялся дефицитом тормозных медиаторов - серотонина и ГАМК [2,3]. ГАМК-А рецептор представляет собой сложный пентамерный комплекс с множеством сайтов связывания позитивных и негативных модуляторов, включая сайты связывания ГАМК, бензодиазепинов, стероидов, алкоголя, барбитуратов, а также блокаторов хлорного ионофора [39]. Роль ГАМК и ГАМК-А рецепторов в патогенезе тревоги у человека и животных хорошо доказана и подробно обсуждается в целом ряде работ [2,35,51]. В целом, подавление ГАМКергической фукнции приводит к тревоге, а активация центральных ГАМК-А рецепторов вызывает анксиолизис [35,50].
Гораздо менее известна роль центральной ГАМК в патогенезе депрессии, хотя имеющиеся данные указывают, что участие ГАМК в механизмах возникновения депрессии очень важно. Впервые ГАМКергическая гипотеза депрессии была сформулирована в 1980 году [19] и за последние десятилетия получила значительное подтверждение в клинике [11,45,67]. В таблице 1 суммированы клинические и экспериментальные данные о влиянии депрессии на ГАМКергическую систему мозга: в целом, у человека и животных наблюдается общее снижение ГАМКергической функции при депрессии и ее восстановление под влиянием различных антидепрессантных факторов [54,67,74]. Таким образом, появляются основания полагать, что именно ГАМКергическая система может участовать как в патогенезе тревоги и депрессии [12,41], так и интеграции данных патологий в единый тревожно-депрессивный континуум (Рис. 1).
Рисунок 1. Роль ГАМК в интеграции тревожно-депрессивной патологии (гипотеза)



![]()
![]()


![]()
![]()

![]()
![]()
![]()
![]()
![]()
Основываясь на данных таблицы 1, следует ожидать, что препараты, стимулирующие ГАМКергическую систему, будут эффективны и при терапии депрессии. В таблице 2 приводятся сведения по некоторым ГАМК-активным АЛ, которые в клинике действительно проявляют выраженное антидепрессантное действие.
Таблица. 1. Влияние депрессии и антидепрессантов на ГАМКергическую систему| Клинические данные | Экспериментальные данные |
| При депрессии | |
| · Снижение уровня ГАМК в плазме, спинномозговой жидкости и мозге у депрессивных больных [12,42,71] | · Снижение уровня ГАМК у крыс в гиппокампе в модели выученной беспомощности [54,67], а также в коре и аккумбенсе в модели вынужденного плавания - тесте Порсолта [11,26]. Повышение плотности ГАМК-А рецепторов в септуме у крыс в модели выученной беспомощности [40]. Повышение связывания мусцимола у крыс, устойчивых в модели поведенческого отчаяния [16] |
| · Снижение уровня ГАМК-активных стероидов и повышение уровня антагонистов стероидов [27,37,38,46, 62,70], в том числе у депрессивных женщин с пре-менструальным синдромом [6] | · Снижение уровня ГАМК-активных стероидов (аллопрегнанолона) в коре в модели социальной изоляции у мышей и крыс [6,62], дисбаланс уровня стероидов в мозге в модели ольфактобульбектомии у крыс [76] |
| · Снижение уровня пептида-ингибитора связывания диазепама в спинно-мозговой жидкости и мозге у депрессивных больных [61,65] и пациентов со смешанной тревогой и депрессией [29] | · Снижение связыванния бензодиазепинов в модели хронического стресса во фронтальной коре, гиппокампе, гипоталамусе; повышение уровня пептида-ингибитора связывания диазепама в мозге в модели хронического стресса у мышей [42] |
| В результате терапии не-ГАМКергическими антидепрессантами | |
| · Повышение уровня ГАМК в коре у депрессивных пациентов [63] | · Повышение уровня ГАМК в гиппокампе у крыс в модели выученной беспомощности [40,54] |
| · Повышение уровня ГАМК в коре у пациентов после электросудорожной терапии [64] | · Повышение уровня ГАМК в стриаутуме, миндалине, гиппокампе и септуме у крыс [26,31] |
| · Восстановление уровня ГАМК-активных стероидных агонистов (аллопрегнанолона) в плазме у депрессивных пациентов [37,38,76]; повышение уровня ГАМК-активных стероидных агонистов и подавление уровня антагонистов в плазме у депрессивных пациентов после выздоровления [62] | · Повышение уровня ГАМК-активных стероидов (аллопрегнанолона) в мозге у крыс [62] |
· Аналоги ГАМК и препараты, влияющие на метаболизм ГАМК:
Тиагабин [67], Бета-фенил-ГАМК [43], Габапентин [24, 28, 67], Аминооксиуксусная кислота [25, 53] (Также оказывает нормотимическое действие [48]), Вигабатрин [8, 60]
· Бензидазепиновые агонисты:
Диазепам, карбамазепин и окскарбазепин [5, 50, 51, 53, 56]
· Этанол [37, 38]· Стероидные агонисты:
Прогестерон, тетрагидропрогестерон, аллопрегланолон [20] (Потенцируют действие других АД [46]).
В подтверждение ГАМКергической теории депрессии у человека, Petty и Sherman [54] впервые указали на роль ГАМК в патогенезе депрессии у животных. Впоследствие эти данные получили дальнейшее подтверждение в работах целого ряда лабораторий с использованием многочисленных животных моделей депрессии [1,40,55]. Таблица 2 суммирует сведения об антидепрессантной активности ГАМК-тропных АЛ при тестировании в различных животных моделях депрессии. Дополняя общую картину, в таблице 3 приведены данные о продепрессантных свойствах ГАМК-тропных анксиогенных препаратов у животных.Таблица 2. Антидепрессантные эффекты ГАМК-активных анксиолитиков в экспериментальных моделях депрессии
Препараты | Модель депрессии и литература |
| Аналоги препараты, влияющие на метаболизм ГАМК Вальпроат натрия Аминооксиуксусная кислота Вигабатрин Гамма-винил-ГАМК Бета-фенил-ГАМК | · Вынужденное плавание (тест Порсолта), мыши [4,21,57]; потенцирует эффекты других АД в модели вынужденного плавания, мыши [73] · Вынужденное плавание, крысы [10,11] · Потенцирует эффекты других АД в модели вынужденного плавания, мыши [73] · Ольфактобульбектомия, крысы [36] · Снимает вызванное стрессом повышение плотности бензодиазепиновых рецепторов в модели вынужденного плавания, мыши [58] |
| Агонисты ГАМК ГАМК Мусцимол | · Вынужденное плавание, мыши [4], выученная беспомощность, крысы [40,67] · Вынужденное плавание, мыши, крысы [4,10, 57]; потенцирует эффекты АД (этанола и аллопрегнанолона) в модели вынужденного плавания, мыши [32,37,38] |
| Бензидазепиновые агонисты Диазепам Карбамазепин и др. | · Вынужденное плавание, крысы [49]; снижает мурицидную агрессию в модели ольфактобульбектомии, крысы [68] · Вынужденное плавание и выученная беспомощность, мыши, крысы [7,34]; потенцируют эффекты других АД в модели вынужденного плавания, мыши [73] |
| Этанол | · Вынужденное плавание, мыши [32]; снижает депрессантное действие кокаина в модели вынужденного плавания, крысы [30] |
| Стероидные агонисты Прогестерон, тетрагидро-прогестерон Аллопрегнанолон | · Вынужденное плавание, крысы; потенцируют эффекты других АД в модели вынужденного плавания, мыши, крысы [20,62] · Вынужденное плавание, мыши [37,62]; потенцирует эффекты других АД в модели вынужденного плавания, мыши [32] |
Таблица 3. Продепрессантные эффекты ГАМК-активных анксиогенных препаратов в экспериментальных моделях депрессии
Препараты | Модель депрессии и литература |
| Антагонисты ГАМК Бикукуллин | · Вынужденное плавание, выученная беспомощность, мыши, крысы [4,30,54], снижает антидепрессантные эффекты вальпроата натрия, этанола, фенгабина и мелатонина в моделях вынужденного плавания и ольфактобульбектомии у крыс и мышей [21,32,44,57] |
| Бензодиазепиновые инверсные агонисты FG 7142, Ro 15-3505 Бета-карболин-3-карбоксилата (БКК) этиловый и метиловый эфиры | · Вынужденное плавание, мыши, крысы [13,69], выученная беспомощность, крысы [74] · Вынужденное плавание, крысы [49], повышенная депрессия в моделях вынужденного плавания и подвешивания за хвост у линии мышей с врожденной повышенной чувствительностью к метиловому эфиру БКК [17] |
| Стероидные антагонисты Прегненолон-сульфат | · Вынужденное плавание, мыши [59] |
| Блокаторы хлорного ионофора Пикротоксин Коразол | · Вынужденное плавание, крысы и мыши [4,69]; снижает антидепрессантные эффекты мусцимола, вальпроата натрия и других АД в моделях вынужденного плавания и выученной беспомощности у крыс [10,21,74] · Вынужденное плавание, крысы [69] |
В целом, сравнительный анализ данных, приводимых в таблицах 1, 2 и 3, позволяет заключить, что у человека и животных активации ГАМКергической системы соответствует снижение тревоги и депрессии, тогда как подавление ГАМКергической активности коррелирует с повышением их уровня. Все это подтверждает важную роль ГАМКергической системы мозга в патогенезе тревоги и депрессии, а также указывает, что поиск новых ГАМК-активных препаратов может быть перспективным направлением при терапии тревоги и депрессии.
Заключение
Важным аспектом в обсуждении ГАМКергической концепции тревоги и депресии является рассмотрение структур мозга, участвующих в патогенезе. Так, хронический стресс – наиболее частая причина депрессии – активирует ГАМКергические области переднего мозга, включая дорзомедиальный гипоталамус и гиппокамп [31] – важные элементы, традиционно относящиеся к депрессогенной системе мозга. Многочисленные работы описывают изменение морфологии гиппокампа при депрессии (см. [66]), указывая на его роль в патогенезе депрессии. Гиппокамп богат ГАМКергическими нейронами и является важным компонентом мозговой ГАМКергической системы. Ведущая роль гиппокампа в механизмах памяти, а также особая роль негативных когниций в патогенезе тревоги и депрессии, может быть одним из механизмов участия ГАМКергической системы мозга в патогенезе тревоги и депрессии [35]. Морфологические и метаболические изменения при депрессии были обнаружены и в миндалине [18,66] – еще одной ГАМКергической структуре мозга, связанной с хранением и обработкой негативной памяти и одновременно участвующей в механизмах тревоги [15]. Подтверждая возможность патогенеза в рамках системы «тревога-депрессия-память-миндалина», Jasnow и Hubman [33] указывают на вовлечение ГАМКергических процессов миндалины в модели депрессии (обусловленное социальное поражение) у грызунов. На важную роль миндалины в интеграции тревоги и депресии указывают и клинические данные с использованием функционального магнитного резонанса [78], отмечающие выраженную активацию миндалины при предъявлении изображений испуганных лиц депрессивным пациентам. Хорошо также известно об участии префронтальной коры в патогенезе различных типов тревоги [14,48]. При этом префронтальная кора традиционно рассматривается как часть «депрессивной» патогенной системы мозга, морфологические и метаболические изменения при депрессии в которой описаны во многих работах [66,78]. Отметим, что ГАМКергические нейроны в ней составляют наибольшую группу клеток. Кроме того, префронтальная кора вовлечена в механизмы памяти и высшие когнитивные функции, что может служить еще одним ГАМКергическим механизмом патогенеза в системе «тревога-депрессия-когниции».
В целом, приводимые нами данные указывают на существенное перекрывание ГАМКергических структур мозга, вовлеченных в патогенез тревоги и депрессии. Это подтверждает наше предположение о существовании в мозге общего ГАМКергического патогенного круга при тревожно-депрессивных расстройствах.
Как уже указывалось ранее, генетические факторы играют большую роль в патогенезе тревоги и депрессии человека и животных [23,48], и поэтому нейрогенетические подходы к пониманию тревожно-депрессивного патогенеза являются чрезвычайно перспективными. Убедительно демонстрируя связь ГАМК и депресии на нейрогенетическом уровне, в двух экспериментальных моделях депрессии (тесте Порсолта и подвешивании за хвост) генетический локус, «ответственный» за депрессивное поведение, был выявлен именно в области, кодирующей одну из субъединиц ГАМК-А рецептора [77] (гены, кодирующие субъединицы ГАМК-А рецепторов, важны при патогенезе тревоги [75]).
Безусловно, помимо ГАМК в мозге существуют и другие механизмы, интегрирующие патогенез тревоги и депрессии. Так, Boyer [9] в статье «Имеют ли тревога и депрессия общий патофизиологический механизм?» указывает на кортиколиберин в качестве одного из таких возможных общих патогенных звеньев. Тем не менее, на фармакологическом, нейрофизиологическом и нейрогенетическом уровне существуют убедительные данные в пользу того, что дисфункции ГАМКергической системы мозга, возможно, являются ведущим интегральным патогенетическим механизмом тревоги и депрессии. Поэтому поиск новых препаратов, воздействующих на многочисленные сайты ГАМКергических рецепторов, может не только привести к появлению новых ГАМК-активных АД и АД, но и способствовать созданию принципиально новых типов психотропных средств. В частности, ГАМКергических препаратов, избирательно эффективных при терапии коморбидности; мнемотропных АЛ/АД-агентов, селективно подавляющих тревогу и депрессию, связанные с негативными когнициями; быстродействующих ГАМК-активных АД; препаратов для терапии смешаной тревоги и депрессии (в том числе неясной симптоматики и генеза); препаратов для купирования острых приступов тревоги и депрессии, в том числе антипанического и антисуицидального действия; АЛ и АД, избирательно действующих в зависимости от пола и др. Поиск дальнейших терапевтических воздействий, направленных на общий патогенетический механизм, может позволить на качественно ином уровне решить проблему терапии тревоги и депресии.
Литература
1. И.В. Белозерцева, Б.В. Андреев, Ж-л ВНД, 47(6), 1024 - 1031 (1997).
2. А.В. Калуев, Эксп. клин. фарм., 60(5), 3 – 7 (1997).
3. А.В. Калуев, Н.А.Нуца, Эксп. клин. фарм., 61(5), 69 – 74 (1998).
4. K.O. Alley, S.K. Kulkarni, Naunyn Schmiedebergs Arch. Pharmacol., 339(3), 306 - 311 (1989).
5. S.V. Argyropoulos, D.J. Nutt, Eur. Neuropsychopharm., 9(6), 407 – 412 (1999).
6. M.L. Barbaccia, S. Lello, T. Sidiropoulou et al., Psychoneuroendocrinol., 25, 659-675 (2000).
7. V. Benjamini, L.L. Skalitz, S.R.L. Joka, R. Andreatini, Eur. J. Pharmacol., 347, 23 – 27 (1998).
8. F.M. Besag, Drug Saf., 24(7), 513 - 536 (2001).
9. P. Boyer, Acta Psych Scand, 406, 24 – 29 (2000).
10. F. Borsini, S. Evangelista, A. Meli, Eur. J. Pharmacol., 121(2), 265 – 268 (1986).
11. F. Borsini, A. Mancinelli, V. D’Arkano, S. Evangelista, A. Meli, Pharmacol. Biochem. Behav., 29(9), 275 – 279 (1988).
12. L. Chang, C.C. Cloak, T. Ernst, J. Clin. Psych., 64(3), 7 – 14 (2003).
13. K. Chopra, J. Kunchandy, S.K. Kulkarni, Arch. Int. Pharmacodyn. Thar., 294, 56 – 63 (1988).
14. R.J. Davidson, Biol. Psych., 51, 68 – 80 (2002).
15. M. Davis, P.J. Whalen, Mol. Psych., 6(1), 13 – 34 (2001).
16. R.C. Drugan, S.M. Paul, J.N. Crawley, Brain Res., 631(2), 270 - 276 (1993).
17. J.C. Do-Rego, C. Saudeau, G. Chapouthier, J. Costentin, Pharmacol. Biochem. Behav., 72, 411 – 416 (2002).
18. W.C. Drevets, Ann. N.Y. Acad. Sci., 877, 614 – 637 (1999).
19. H.M. Emrich, D.V. Zervessen, W. Kissling et al., Arch. Psych. Nervenkr., 229, 1-16 (1980).
20. E. Estrada-Camarena, C.M. Contreras, M. Saavedra et al., Behav. Brain Res., 134, 175 – 183 (2002).
21. A. Fernandes Teruel, F. Boix, P. Escorihuela et al., Eur. J. Pharmacol., 152 (1-2), 1 - 7 (1988).
22. M. Freeman, S.A. Freeman, S.L. McElroy, J. Affect. Disorders, 68, 1 – 23 (2002).
23. D.A. Finn, M.T. Rutledge-Gorman, J.C. Crabbe, Neurogenet., 4, 109 – 135 (2003).
24. G.C.Y. Fong, B.M.Y. Cheung, C.R. Kurama, Med. Prog., 2, 1 – 8 (2003).
25. W.S. Gilmer, Expert Opin. Pharmacother., 2(10), 1597 – 1608 (2001).
26. R. Gomez, C.R. Vargas, M. Wajner, H.M.T. Barros, Brain Res., 968, 281 – 284 (2003).
27. M.S. George, A. Guidotti, D. Rubinov et al., Biol. Psych., 35 (10), 775 – 780 (1994).
28. S.N. Ghaemi, J.J. Katzow, S.P. Desai, F.K. Goodwin, J. Clin. Psych., 59 (8), 426 – 429 (1998).
29. A. Guidotti, Neuropharmacol., 30 (12), 1425 – 1433 (1991).
30. T. Hayase, Y. Yamamoto, K. Yamamoto, BMC Pharmacol., 2, 1 – 9 (2002).
31. J.P. Herman, A. Renda, B. Bodie, Biol. Psych., 53, 166 – 174 (2003).
32. K. Hirani, R.T. Khisti., C.T. Chopde, Neuropharmacol., 43, 1339 – 1350 (2002).
33. A.M. Jasnow, K.L. Hubman, Brain Res., 920 (1-2), 142 – 150 (2001).
34. S.R.L. Joka, L.L. Scalitz, V. Benjamini et al., Eur. Neuropsychopharmacol., 10, 223 – 228 (2003).
35. A.V. Kalueff, D.J. Nutt, Depression Anxiety, 4, 100 – 110 (1996-1997).
36. J.P. Kelly, A.S. Wrynn, B.E. Leonard, Pharmacol. Ther., 74(3), 299 – 316 (1997).
37. R.T. Khisti, S.N. Penland, M.J. VanDoren et al, World J. Biol. Psych., 3, 87 – 95 (2000).
38. R.T. Khisti, C.T. Chopde, S.P. Jain, Pharmacol. Biochem. Behav., 67, 137 - 143 (2000).
39. E.R. Korpi, G. Grunder, H. Luddens, Prog. Neurobiol., 67, 113 – 159 (2003).
40. M.L. Kram, G.L. Kramer, M. Steciuk et al., Neurosci Res., 38, 193 – 198 (2000).
41. J.H. Krystal, G. Sanacora, H. Blumberg et al., Mol. Psych., 7, 71 – 80 (2002).
42. A. Kugaya, G. Sanacora, N.P.L.G. Verhoeff et al., Biol. Psych., 53, 1 – 8, (2003).
43. I.P. Lapin, CNS Drug Dev., 7(4), 471 - 481 (2001).
44. K.G. Lloyd, B. Zibkovic, D. Sanger et al., J. Pharmacol. Exp. Ther., 241(1), 245 – 250 (1987).
45. K.G. Lloyd, B. Zibkovic, B. Skatton et al., Prog. Neuropsychopharmacol. Biol. Psych., 13(3-4), 341 – 351 (1989).
46. R.B. Lydiard, J. Clin. Psych., 64, 1 – 7 (2003).
47. R. Nesse, Neurosсi Biobehav. Revs, 23, 895 - 903 (1999).
48. E.J. Nestler, E. Gould, H. Manji et al., Biol. Psych., 52, 503 – 528 (2002).
49. H. Nishimura, Y. Ida, A. Tsuda, M. Tanaka, Pharmacol. Biochem. Behav., 33 (1), 227 – 231 (1989).
50. D. Nutt, Understanding Mental Health, Octopus Ltd, Bristol (2001), 1 - 3.
51. D.Nutt, Eur. Neuropsychopharm., 10(4), 433 – 437 (2001).
52. D.J. Nutt, J.C. Ballender, D.Sheehan, H.U. Whittchen, Int. J. Neuropsychopharmacol., 5, 315 – 325 (2002).
53. G. Obrocea, R.M. Dunn, M.A. Frye et al., Biol. Psych., 51, 253 – 260 (2002).
54. F. Petty, A.D. Sherman, Pharmacol. Biochem. Behav., 18(4), 649-50 (1981).
55. F. Petty, J. Affect. Disorders, 34, 275 – 281 (1995).
56. R.M. Post, T.A. Ketter, K. Denicoff et al., Psychopharmacol., 128(2), 115 – 129 (1996).
57. V. Raghavendra, G. Kaur, S.K. Kulkarni, Eur. Neuropsychopharmacol., 10, 473 – 481 (2000).
58. D. Reddy, TINS, 24(3), 103 – 106 (2001).
59. L. Rago, R.A. Kiivet, A. Adojaan et al., Pharmacol. Toxicol., 66(1), 41 – 44 (1990).
60. H.A. Ring, R. Crellin, S. Kirker, E.H. Reynolds, J. Neurol. Neurosurg. Psych., 56(8), 925 - 928 (1993).
61. A. Roy, Neuropharmacol., 30 (12), 1441 – 1444 (1991).
62. R. Rupprecht, Psychoneuroendocrinol., 28, 139 – 168 (2003).
63. G. Sanacora, G.F. Mason, D.L. Rothman, J.H. Krystal, Am. J. Psych., 154(4), 663 – 665 (2002).
64. G. Sanacora, G.F. Mason, D.L. Rothman et al., Am. J. Psych., 160(3), 577 – 579 (2003).
65. J.J. Sandford, S.V. Argyropoulos, D.J. Nutt, Pharmacol. Ther., 88, 197 – 227 (2000).
66. Y. Sheline, Biol. Psych., 54, 338 – 352 (2003).
67. I.S. Shiah, L.N. Yatham., Y.C. Gau, G.B. Baker, Prog. Neuropsychopharmacol. Biol. Psych., 27, 419 – 423 (1998).
68. S. Shibata, H. Nakanishi, S. Watanabe, S. Ueki, Pharmacol. Biochem. Behav., 21(2), 225 – 230 (1984).
69. T. Skrebuhova, L. Alikmats, V. Matto, Meth. Find. Exp. Clin. Pharmacol., 21(3), 173 – 178 (1999).
70. B. Spivak, R. Maayan, M. Kotler et al., Psychol. Med., 30(5), 1227 – 1231 (2000).
71. I. Sundman-Erikkson, P. Alard, J. Affect. Disorders, 71, 29 – 33 (2002).
72. I. Sundstrom Poromaa, S. Smith, M. Gulinello, Arch. Women Ment. Health, 6, 23 - 41 (2003).
73. G. Szymczyk, I. Zebrovska-Lupina, Pol. J. Pharmacol., 52(5), 337 – 344 (2000).
74. G. Tunnicliff, E. Malatynska, Neurochem. Res., 28(6), 965 – 976 (2003).
75. O.Yu. Vekovisheva, Academic Dissertation, Tampere (2003).
76. M. Uzunova, M. Ceci, C. Kohler et al., Brain Res., 976, 1 – 8 (2003).
77. T. Yoshikawa, A. Watanabe, Y. Ishitsuka et al., Genome Res., 4, 357 – 366 (2002).
78. D.A. Yurgelun-Todd, S.A. Gruber, G. Kanayama et al., Bipolar Disorders, 2(3), 237 – 248 (2000).
79. Вестник биологической психиатрии №12, 2003г. – электронный бюллетень РОБП и УОБП.
Похожие работы




... . Начался поиск принципиально новых решений. 1.2 Ноотропил и другие лекарственные средства В начале 60-х годов бельгийские фармакологи получили соединение, которое представляло собой гамма-аминомасляную кислоту, свернутую в кольцо и снабженную некоторыми дополнительными радикалами. Профессор К. Джурджеа и его сотрудники установили, что это соединение значительно улучшает память и облегчает ...

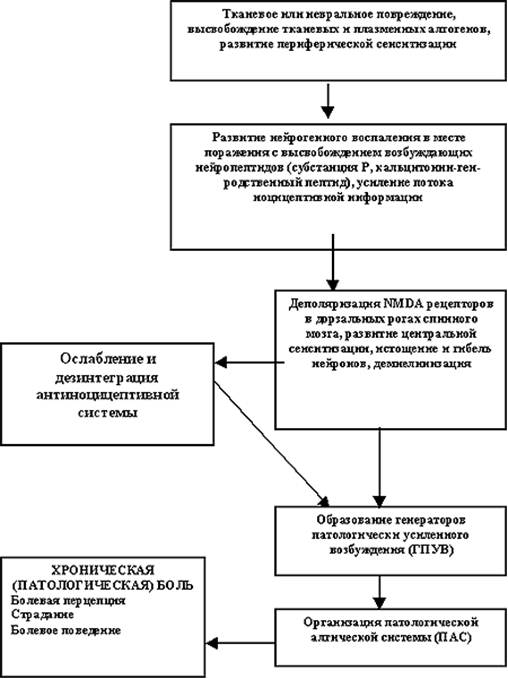
... . 6. Формирование патологической алгической системы (ПАС), включающей различные уровни центральной нервной системы и определяющей течение и характер всех компонентов патологической боли: болевой перцепции, страдания и болевого поведения. Хроническая боль как общепатологическая проблема Что же представляет собой хроническая (патологическая) боль с позиций общей патологии? В учении о болезни ...
"приказы", обратные по своей направленности: например, сосудам и зрачку - сузиться, мускулатуре мочевого пузыря - сократиться и так далее. Вегетативная нервная система очень чувствительна к эмоциональному воздействию. Печаль, гнев, тревога, страх, апатия, половое возбуждение - эти чувства вызывают изменения функций органов, находящихся под контролем вегетативной нервной системы. Например, ...
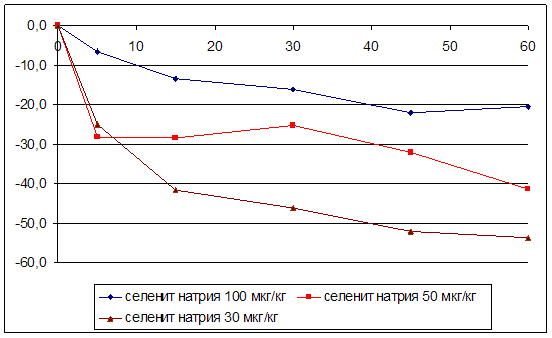
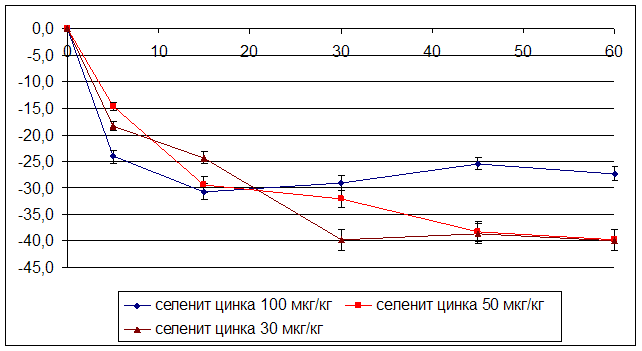
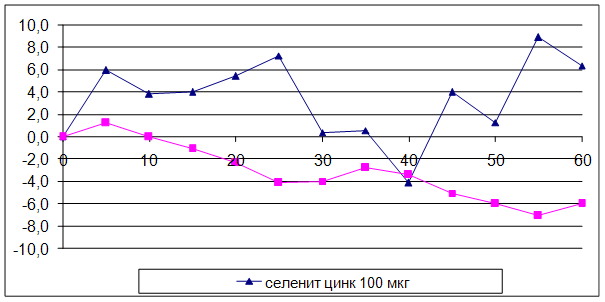
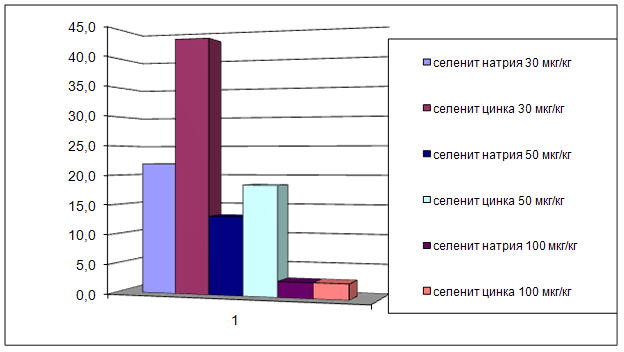
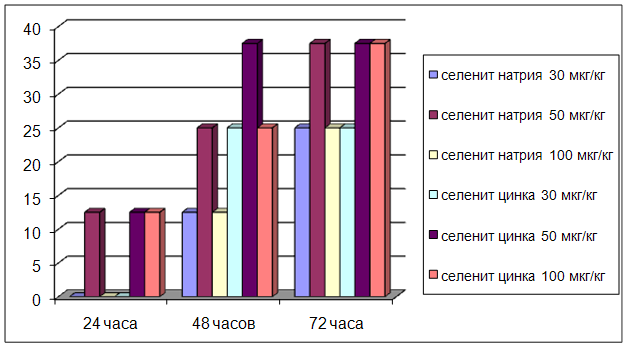
... на мозговой кровоток, динамику развития постишемических феноменов и психоневрологический статус остаётся неизученным. Указанные обстоятельства и послужили основанием для сравнительного изучения влияния селенита натрия и селенита цинка на системную гемодинамику и мозговой кровоток в условиях нормы и при патологических состояниях. ГЛАВА 2 МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ 2.1 Изучение острой ...
0 комментариев