Физиологическое значение текучести мембран
Спиновые метки, использующиеся при изучении текучести мембран
Вращение мембранных белков
Латеральная диффузия липидов и белков в мембранах
Примеры латеральной диффузии компонентов мембран
Липидно-белковые взаимодействия
Изменения в липидном бислое, связанные с присутствием интегральных мембранных белков
Динамические свойства остова мембранных белков и их боковых цепей
Навигация
Спиновые метки, использующиеся при изучении текучести мембран
Динамическое поведение мембранных систем и липидно-белковые взаимодействия
70319
знаков
0
таблиц
8
изображений
2.2 Спиновые метки, использующиеся при изучении текучести мембран
ЭПР — это очень ценный метод изучения мембран. Обычно в качестве меток используются стабильные парамагнитные соединения, содержащие нитроксидный радикал. Неспаренный электрон при наложении сильного магнитного поля переходит с одного энергетического уровня на другой под действием микроволнового излучения. Метод очень чувствителен: обычно спектр регистрируется при концентрации спиновых меток около 10"*М в 50 мкл образца. Спектр ЭПР, как правило, представляют в виде первой производной от спектра поглощения.
В спектре нитроксидного радикала имеются три пика, отвечающие спин-спиновым взаимодействиям неспаренного электрона и ядра атома азота. И положение спектра, и расщепление, обусловленное спином ядра, зависят от ориентации молекулы относительно внешнего поля. Как мы уже говорили в предыдущем разделе, это означает, что характер спектра зависит от характера вращения молекулы. На рис. 5.3 приведены спектры для самых разных случаев — от свободного вращения до полной неподвижности. Для того чтобы найти тк в предположении, что вращательное движение молекулы изотропно, можно использовать простое уравнение, в которое входит высота пиков. Этот подход наиболее применим для такой метки, как TEMPO.
Спиновые метки — производные жирных кислот или фосфолипидов, — конечно, не вращаются изотропно, и в этом случае для нахождения параметра упорядоченности S из величины расщепления между линиями спектра используется вторая простая формула. Предположив, что такие метки имеют форму жесткого стержня, можно из величины параметра S оценить максимальное отклонение зонда от нормали к поверхности мембраны.
Многие предположения, которые делаются при нахождении из спектров ЭПР времен вращательной корреляции или параметра S, снижают ценность этого метода для определения количественных характеристик молекул. Эти параметры тем не менее позволяют получить качественное представление о поведении мембраны. Результаты измерения степени упорядоченности спиновых меток, фиксированных в мембране на разной глубине, свидетельствуют об увеличении неупорядоченности в направлении от поверхности мембраны к ее центральной области. Качественно такая же картина получена с помощью 2Н-ЯМР. Во многих исследованиях проводилось сравнение текучести мембран, которую определяли по данным о величинах тк и S для спиновых меток, при различных возмущающих воздействиях.
2.3 Флуоресцентные зонды, используемые для измерения текучести мембран
Для измерения вращательной диффузии молекул давно используется метод деполяризации флуоресценции. Вращение некоторых зондов в мембранах часто сравнивают с их вращением в маслах с известной вязкостью, при этом пользуются понятием «микровязкость», которую измеряют в пуазах. Термин «микровязкость» означает, что зонд «чувствует» величину вязкости только в своем непосредственном окружении, т. е. этот параметр не является макроскопическим. С точки зрения молекулярного движения подобные измерения не имеют строгого количественного смысла, но, как и в случае со спиновыми метками, полученные результаты позволяют создать ценную качественную картину.
В табл. 1 приведены структурные формулы некоторых зондов, используемых при измерении текучести мембран. Чаще всего применяют ДФГ, поскольку он с легкостью накапливается в мембранах, обладает интенсивной флуоресценцией, по-видимому, не связывается с белками и чувствителен к физическому состоянию мембран. Однако временные измерения свидетельствуют о некой гетерогенности неизвестной природы; кроме того, имеются разногласия по поводу ориентации зонда в бислое и характера его подвижности.
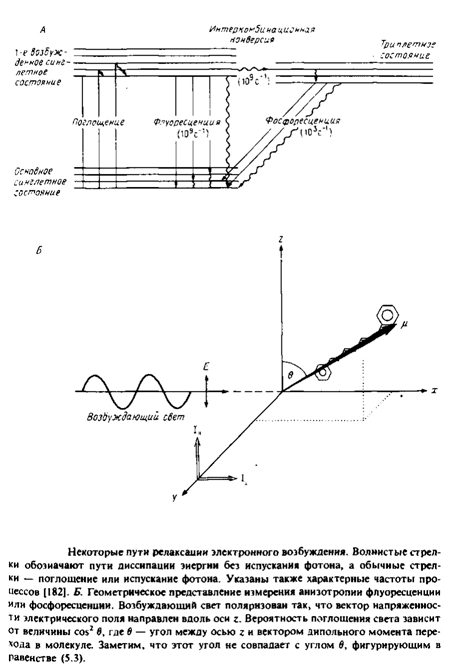
Рис. 4 иллюстрирует принципы, лежащие в основе применения флуоресценции и фосфоресценции для анализа вращения молекул. Молекула, поглотившая фотон, за время 10" 11 с переходит в возбужденное синглетное состояние Si. Термин «синглетное» указывает на то, что спин электрона при поглощении фотона не изменяется. Обычно возвращение в основное, невозбужденное состояние происходит за время - 10"8 с. Причинами такого перехода являются столкновение с соседними молекулами, диссипация энергии в виде тепла или испускание фотона. Характерное время перехода популяции молекул из синглетного возбужденного состояния в основное равно тт. Некоторые молекулы с большой вероятностью переходят из синглетного возбужденного состояния в триплетное, и этот переход сопровождается изменением спина электрона. Испускание фотона при переходе молекулы из этого состояния в основное называется фосфоресценцией. Время жизни молекулы в триплетном состоянии относительно велико, что связано с изменением спинового состояния электрона при испускании фотона.
Рассмотрим принцип метода деполяризации. Свет, поляризованный вдоль оси z, будет предпочтительно возбуждать молекулы, у которых дипольный момент перехода имеет такую же ориентацию. Вероятность возбуждения пропорциональна cos2 в, где в — угол между дипольиым моментом перехода и вектором напряженности электрического поля. Такая же зависимость от cos2 в имеет место и для процесса испускания фотона, поэтому молекула, у которой дипольный момент перехода ориентирован вдоль оси z, будет испускать свет с такой же поляризацией.
Испускаемый свет анализируют с помощью поляризаторов, определяя интенсивность компоненты, параллельной и перпендикулярной направлению поляризации возбуждающего света.
Анизотропия г связана с /, и IL следующим образом:

Если между моментами поглощения и испускания света молекула не вращается, то /, будет больше, чем 1±, и г будет иметь максимальное значение 0.4. Если же молекулы, находясь в возбужденном состоянии, совершают интенсивные изотропные вращательные движения, то г будет равно нулю, поскольку в среднем молекулы к моменту испускания света будут ориентированы беспорядочно. При тк = tf величина г будет зависеть от характера движения. При этом т ~ 1 и ту ' отражают скорости вращения и испускания света.
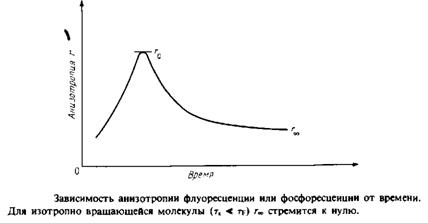
Для того чтобы разграничить динамические эффекты и эффекты, связанные с ограничениями, налагаемыми на диапазон допустимых движений, определяют временную зависимость анизотропии r света, испускаемого молекулой после возбуждения. Значение г в данный момент времени отражает степень разу-порядочивания молекул относительно вектора поляризации за вре- ■ мя, прошедшее после поглощения возбуждающего импульса света. Спад r от максимального значения го задается выражением
![]()
тк — время вращательной корреляции, оно определяется скоростью затухания. Если при / оо все молекулы окажутся беспорядочно ориентированными, то л в пределе будет равно нулю. Однако если движение молекул ограничено, то их ориентация никогда не будет совершенно случайной. В такой ситуации г» не будет равно нулю, и с помощью этого параметра можно определить степень ограничения, которое налагается на подвижность зонда его окружением. Параметр упорядоченности определяется из этих величин так же, как в случае ЭПР или ЯМР:
![]()
К сожалению, такие кинетические измерения проводятся редко, большинство экспериментов выполнено в стационарном режиме при непрерывном возбуждении и испускании. Значение г, полученное таким способом, носит усредненный характер и определяется как
![]()
Рассмотрим следующие три случая.
1. Очень быстрое движение: 7"= г„, предельному значению.
2. Очень медленное движение или его отсутствие: г = го, максимальному значению.
3. Случаи, представляющие практический интерес '- 7 зависит как от скорости движений, так и от ограничений, налагаемых на них.
Без соответствующей корректировки определение микровязкости по результатам измерений 7 не будет сводиться к простым динамическим измерениям.
Вязкость бислоя весьма высока, поэтому время вращательной корреляции небольших флуоресцентных и ЭПР-зондов составляет 10~8—10 с. В воде, имеющей вязкость 0,01 пуаз, молекулы такого размера вращались бы по крайней мере в 100 раз быстрее. Мембранные белки имеют значительно большие размеры, чем упомянутые метки, и поэтому вращаются гораздо медленнее. Чтобы метки, «пришитые» к этим белкам, были чувствительны к их вращению, время жизни меток в возбужденном состоянии должно составлять порядка 10~3 с.
Похожие работы
... является курсом, для изучения которого необходимо наличие знаний об основных принципах организации биологических молекул, строении и механизмах действия ферментов. Дисциплина биохимия мембран относится к дисциплинам специализации федерального компонента. 5. Распределение времени, отведенного на изучение дисциплины по учебному плану Форма учебной работы Форма обучения Очная По ...
... Рисунок Кинетика биопроцессов Динамические свойства биопроцессов Каждая система состоящая из элементов будет характеризоваться динамикой, складывающейся из элементов. Кинетика биопроцессов – раздел биофизики, изучающий динамические свойства биопроцессов. 1. Параметры, меняющие свое значение со временем. Переменные величины: численность клеток, биомасса, концентрация ...



... и индуцируют трансмембранную миграцию липидных молекул, поэтому полученные результаты бывает трудно интерпретировать. Детальная оценка достоинств и недостатков методов изучения липидной асимметрии в мембранах дается, например, в обзорах. МЕТОДЫ УСТАНОВЛЕНИЯ ТРАНСМЕМБРАННОГО РАСПРЕДЕЛЕНИЯ ЛИПИДОВ Химическая модификация фосфолипидов Относительно легко подвергаются химической модификации только ...
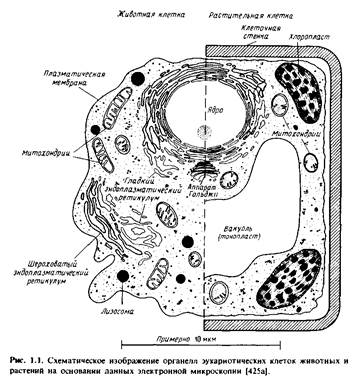
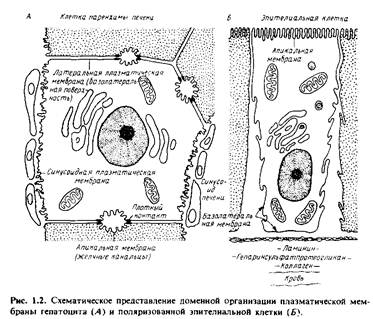
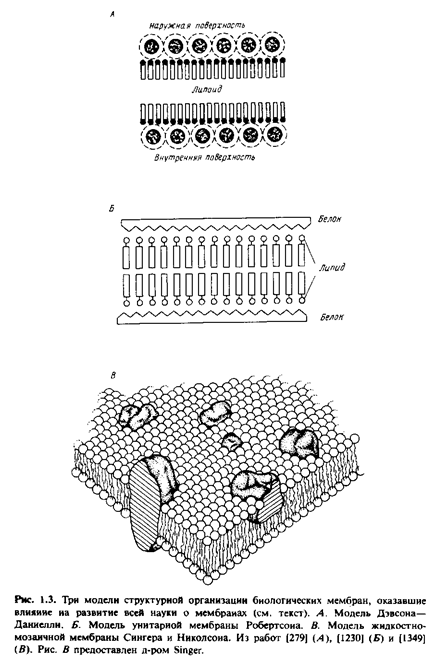
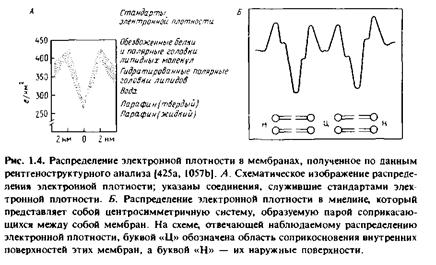
... правильным только благодаря взаимной компенсации ошибок, однако в историческом плане эта работа имела большое значение, поскольку с тех пор концепция липидного бислоя как структурной основы биологических мембран стала доминирующей и на самом деле оказалась верной. Концепция бимолекулярной липидной мембраны получила дальнейшее развитие в предложенной в 1935 г. модели Дэвсона-Даниелли, или ...
0 комментариев