Навигация
Трансляция мРНК у эукариот
85334
знака
0
таблиц
0
изображений
8. Трансляция мРНК у эукариот
Процесс трансляции эукариотической мРНК в основном аналогичен таковому прокариотической мРНК. За некоторыми отмеченными выше исключениями, генетический код универсален и кодоны последовательно транслируются с помощью специфических аминоацил-тРНК-синтетаз на рибосомах. Есть, однако, и три явных различия, обусловленных определенными свойствами эукариотических клеток. Во-первых, аппараты транскрипции и трансляции у эукариот физически разобщены, поскольку транскрипция осуществляется в ядре, а трансляция – в цитоплазме. Во-вторых, на 5'- и 3'-концах эукариотических мРНК имеются особые структуры. И в-третьих, эукариотические мРНК, за исключением мРНК, транскрибируемых с ДНК геномов вирусов, обычно содержат только одну белок-кодирующую последовательность.
Структура и свойства участников трансляции эукариотической мРНК пока изучены гораздо хуже, чем у прокариот. И хотя у эукариот выделяют те же три стадии процесса – инициацию, элонгацию и терминацию, – на каждой из них требуется больше нерибосомных белковых факторов. Несмотря на эти различия, последовательности, кодирующие белки прокариот, нормально транслируются эукариотическими системами трансляции при условии соответствующей модификации их мРНК на 3'- и 5'-концах. И наоборот, кодирующие последовательности эукариот эффективно транслируются системами прокариот, если у них перед 5'-концом инициаторного кодона AUG имеется последовательность Шайна-Дальгарно. Это значит, что трансляционные аппараты обоих типов организмов могут осуществлять свои функции, несмотря на особенности нуклеотидных последовательностей мРНК из разных источников.
а. Особые модификации мРНК эукариот
У эукариотических мРНК, транскрибированных с ядерных или вирусных геномов РНК-полимеразой II, всегда модифицированы 5'-концы, которые в этом случае называют «кэпами». РНК, транскрибируемые эукариотическими РНК-полимеразами I и III, не кэпированы и имеют обычные 5'-фосфатные концы. У большинства мРНК, синтезируемых РНК-содержащими вирусами животных, также имеются кэпы, хотя они синтезируются вирусными РНК-транскриптазами. Многие некэпированные мРНК неэффективно транслируются эукариотическими белоксинтезирующими системами из-за слабого связывания рибосом с мРНК. Кэпирование происходит на 5'-нуклеозидтрифосфате вскоре после инициации синтеза РНК-транскриптов и задолго до его завершения.
На 3'-концах эукариотических мРНК имеются еще и полиаденилатные последовательности. Такой 3' – «хвост» из 50–200 аденилатных остатков не кодируется смысловыми последовательностями соответствующих генов, а присоединяется посттранскрипционно, после разрезания транскрипта в специфическом месте за сигналом терминации трансляции.
б. Инициация трансляции на 5'-кэпированных концах малыми рибосомными субчастицами
Как мы уже говорили, обязательным этапом при инициации трансляции прокариотической мРНК является диссоциация 70S-рибосомы; точно так же и 80S-рибосомы должны диссоциировать до начала трансляции эукариотических мРНК. Малая субчастица в комплексе со множеством белков-помощников, из которых один или несколько нужны для диссоциации рибосомы на составляющие субчастицы, связывает особую инициаторную met-тPHK. И вновь для связывания инициаторной аминоацил-тРНК необходимы GTP и особый белок – eIF-2. У эукариот, однако, met-TPHK не подвергается N-формилированию; но, как и у прокариот, структура тРНК отличается от структуры тPHKMMet. Комплекс 40S, содержащий met-тPHKIMet, GTP и целую армию других факторов eIF, связывается с мРНК вблизи кэпированного 5'-конца или прямо с этим концом; по крайней мере один из этих факторов узнает кэп и связывается с ним. Малая субчастица перемещается при помощи неизвестного пока механизма от кэпированного конца до первого кодона AUG. Никакой последовательности вблизи кодона AUG, аналогичной последовательности Шайна-Дальгарно, пока не обнаружено. Однако эффективность работы AUG в качестве инициатор-ного кодона зависит от наличия определенных фланкирующих нуклеотидов. Такой преинициаторный комплекс объединяется с 60S-субчастицей в ходе энерго- и фактор-зависимой реакции с образованием функционального комплекса инициации. Инициация белкового синтеза может регулироваться фосфорилированием и дефосфорилированием eIF-2.
в. Элонгация и терминация полипептидной цепи
Поэтапная трансляция последовательных кодонов с помощью аминоацил-тРНК у эу- и прокариот в принципе сходна. GTP и фактор элонгации eEF-1, соответствующий прокариотическому комплексу EF-Tu и EF-Ts, периодически поставляют рибосомам аминоацил-тРНК. GTP и eEF-2, являющиеся функциональными аналогами прокариотического EF-G, осуществляют транслокацию. Терминация трансляции у эукариот также происходит в одном из трех стоп-кодонов и сопровождается отделением свободных полипептидных цепей, мРНК и, возможно, 80S-рибосом от мРНК. По-видимому, всю серию терминационных событий осуществляют один фактор eRF и GTP.
В результате последовательных актов инициации образуются полисомы и одновременно, как и у прокариот, начинается трансляция множества кодирующих белки последовательностей. Заметное отличие трансляции у эукариот состоит в том, что клеточные мРНК обычно содержат одну-единственную кодирующую последовательность. Если в мРНК имеется несколько таких областей, то последующие либо вовсе не транслируются, либо транслируются неэффективно.
Кроме регуляции на уровне трансляции при образовании рибосомных белков осуществляется также регуляция по типу обратной связи при транскрипции оперонов рибосомных белков. Мы не будем обсуждать регуляцию этих оперонов на уровне транскрипции, а лишь отметим, что могут происходить репрессия и аттенуация транскрипции рибосомными субчастицами или даже целыми рибосомами. Наиболее вероятно, однако, что ключевым пунктом в регуляции скорости синтеза рибосом является образование рРНК. Таким образом, синтез рибосомных белков и тем самым сборка рибосом регулируются путем изменения содержания рРНК.
Похожие работы
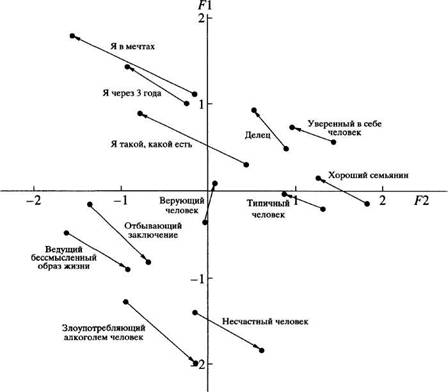
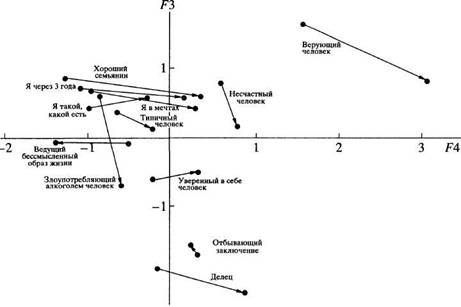
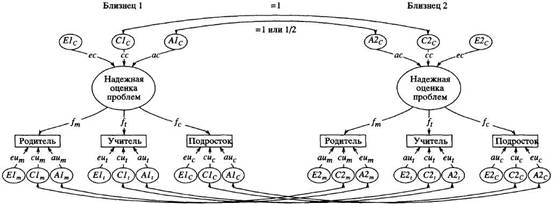
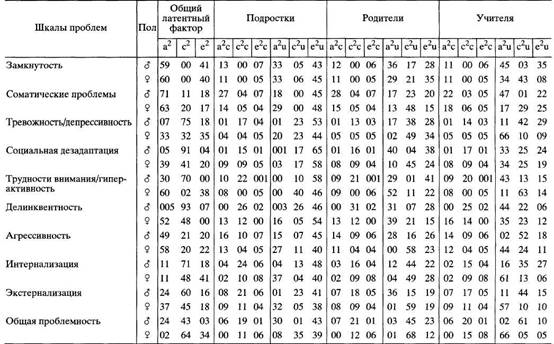
... в отечественной теории и практике психологических измерений. Хотя концепт осмысленности измерения развивается с трансформацией идей Стивенса и разработкой проблем статистики и логики, его положения относительно шкалирования, по проблемам измерений в психологии и связанной с ними осмысленностью измерений требуют, на наш взгляд, критического анализа привычной практики использования психологического ...
... , вызванные динамическими му-тациями.-----------------------T-----------T-------T-----T------T------T----------------------¬ Болезнь, номер по ¦ Ген, лока-¦Триплет¦Норма¦Прему-¦Мута- ¦Литература ¦ МакКьюсику (MIM) ¦ лизация ¦ ¦ ¦тация ¦ция ¦ ¦ -----------------------+-----------+-------+-----+------+------+----------------------+ Синдром ломкой X-хро- ¦FMR1, FRAXA¦(CGG)n ...

... результаты были получены благодаря широкому использованию в лабораторных тестах генетически охарактеризованных животных, а также применению основных методов анализа генетических различий. В настоящее время исследование когнитивных способностей животных в этом тесте является одним из ведущих подходов в оценке особенностей поведения трансгенных животных и мышей-нокаутов. Детальнее с этими вопросами ...
... к основному тону, к тонике. Из обостренности интонации вводного тона среди полутонов развивается еще ряд важных для формы, как процесса интонирования, следствий: в отношении ладовом стирается тетрахордность, давнее наследие средиземноморской музыкальной культуры, и звукоряд европейский становится интонационным единством — гаммой; отношении эволюции гармонии возникает чувство доминантности, все ...
0 комментариев