Навигация
3.2 Выделение хромосом
В ходе описываемых процедур для предотвращения потерь и поломок хромосом, для обеспечения температурного режима необходимо использовать пластиковые пипетки и пробирки. Клетки блокируют на стадии митоза, митотические хромосомы высвобождают воздействием гипотонического шока и гомогенизацией. Хромосомы очищают дифференциальным центрифугированием.
Таблица. Исходные растворы на CMGT
| Среда для роста клеток и селективная среда | NB. Трансфекцию необходимо проводить в средах с небольшим содержанием фосфатов, таких, как DMEM. Клетки, растущие в средах, обогащенных фосфатами, непосредственно перед проведением трансфекции пересейте на среду, бедную фосфатами. Среды, обогащенные фосфатами, можно использовать при глицериновом шоке |
| Гипотонический раствор | 10 мМ Hepes 3 мМ хлорида кальция |
| Раствор для трансфекции | 25 мМ Hepes 134 мМ хлорида натрия 5 мМ хлорида калия 0,7 мМ дигидрофосфата натрия 5 мМ глюкозы |
| 1,25 М хлорида кальция Раствор для отмывки | 25 мМ Hepes 134 мМ хлорида натрия 5 мМ хлорида калия 0,7 мМ дигидрофосфата натрия |
3.3 Перенос хромосом
Процесс переноса хромосом в этом случае очень напоминает описываемый в методе DMGT. Хромосомы осаждают на поверхности клеток хлоридом кальция, и спустя несколько часов клетки обрабатывают реагентом, способным перфорировать мембраны. Здесь тоже важно использовать пластиковые пробирки и пипетки. Последовательность действий, которая приведена ниже, разработана Нельсоном.
1. За день перед проведением трансфекции высейте по 5Х Х105 клеток на 10 чашек диаметром 9 см. Используйте низкофосфатную среду, например DMEM.
2. Ресуспендируйте 108 хромосом в 9 мл раствора для трансфекции.
3. Медленно добавьте к хромосомам 1 мл 1,25 М раствора СаС12, одновременно продувая воздух через суспензию хромосом.
4. Инкубируйте 20–30 мин при комнатной температуре для: образования смеси фосфат кальция – ДНК.
5. Добавьте по 1 мл этой смеси к среде в каждую чашку с реципиентными клетками. Инкубируйте клетки с хромосомами 4–6 ч в увлажняющем инкубаторе при 37 °С.
6. Удалите среду и добавьте 10 мл отмывочного раствора.
7. Удалите отмывочный раствор и обработайте клетки 1 мл среды для глицеринового шока в течение 4 мин при комнатной температуре.
8. Отмойте клетки 3 раза промывочным раствором и инкубируйте в течение ночи в неселективной среде для роста клеток.
9. Через 24 ч поменяйте среду на селективную. Меняйте среду на свежую каждые 3–4 дня.
10. Колонии появятся на 14–21-й день.
3.4 Предварительная селекция
При использовании DMGT донорная геномная ДНК обычно переносится вместе с плазмидой, кодирующей доминантный селективный маркер. Предварительная селекция, выявляющая включение плазмидной ДНК, позволяет получить 100-кратное обогащение клетками, содержащими интересующий нас клеточный ген. Аналогичный прием может быть использован и в CMGT. Для проведения котрансфекции необходимое количество плазмидной ДНК добавляют к суспензии хромосом перед преципитацией хлоридом кальция. Обычно мы добавляем плазмидную ДНК в количестве, достаточном для достижения соотношения 20:1. Селекцию проводим спутся 24 ч после хромосомной трансфекции.
3.5 Возможные ошибки и варианты методики
В литературе описано множество методов выделения хромосом из клеток, блокированных в метафазе. Процедуры очистки тоже разнообразны. Одни из них позволяют получить высокоочищенные препараты, другие–грубую фракцию хромосом, загрязненную разными компонентами клетки. Мы предпочитаем использовать для проведения трансфекции именно такие грубые препараты, во-первых, потому что их получение занимает мало времени, а во-вторых, потому что хромосомы при этом оказываются наименее разрушенными.
Анализ, проведенный Льюисом, показал, что существует линейная зависимость частоты CMGT-трансфекции от дозы донорных хромосом. В большинстве последующих экспериментов исследователи старались ввести в клетку как можно больше хромосом. Однако на практике количество хромосом, которое можно получить, ограничено. Основное препятствие в использовании очень большого количества донорных клеток – это высокая вязкость суспензии хромосом, которая способствует их агглютинации. Вот почему мы добавляем не более 20 хромосом на одну реципиентную клетку. Помимо механического воздействия для получения препарата хромосом можно применять и химическую обработку, включая использование мягких детергентов, таких, как дигитонин.
Исходя из нашего опыта, можно заключить, что результаты трансфекции воспроизводимы. В некоторых случаях может оказаться необходимым оптимизировать условия «шока», варьируя концентрацию глицерина и время инкубации. Обсуждение способа трансфекции с помощью осаждения фосфатом кальций приводится в разд. 6.
При использовании метода CM.GT образуются реципиентные клетки, содержащие фрагменты донорных хромосом: в некоторых случаях они встраиваются в геном реципиента, иногда реплицируются самостоятельно. Невозможно выделить параметр, контролирующий размеры передаваемого фрагмента, и в большинстве экспериментов получаются клоны, содержащие донорный материал в широком диапазоне. Мы детально анализировали введенные фрагменты во всех случаях. В них наблюдались перестройки: это либо внутренние делеции, либо переобогащение альфоидными последовательностями из области центромеры. Внутренние делеции описаны также другими авторами.
Похожие работы
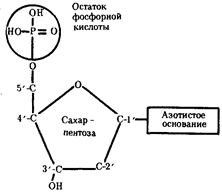
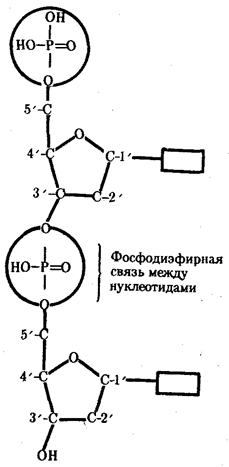
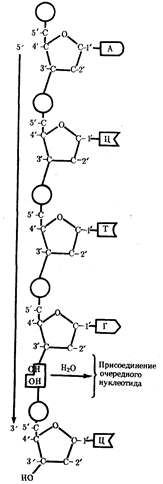
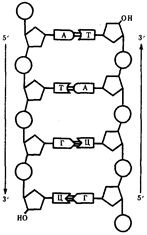
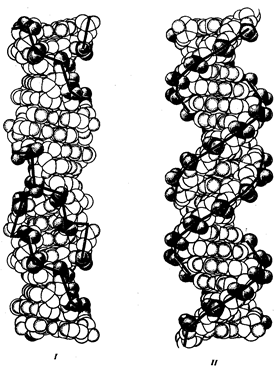
... : генный, хромосомный и геномный. На каждом из них проявляются основные свойства материала наследственности и изменчивости и определенные закономерности его передачи и функционирования. 4. Генный уровень организации генетического аппарата Элементарной функциональной единицей генетического аппарата, определяющей возможность развития отдельного признака клетки или организма данного вида, ...
... , доступные генетическому анализу — это прежде всего скорость и полнота лизиса инфицированных клеток и круг бактерий-хозяев, поражаемых фагами. Широкое распространение в генетическом анализе бактериофагов получили мутанты с условным проявлением. Это мутанты, чувствительные к повышению и понижению температуры, — так называемые термочувствительные (ts) и холодочувствительные (cs). Они нормально ...
... 1972 год, когда в Стенфордском университете П. Берг и С. Коэн с сотрудниками создали первую рекомбинантную ДНК, содержавшую фрагменты ДНК вируса SV40, бактериофага и E. coli. Генетическая инженерия Одним из разделов молекулярной генетики и молекулярной биологии, который нашел наибольшее практическое приложение, является генная инженерия. Генная инженерия – это сумма методов, позволяющих ...
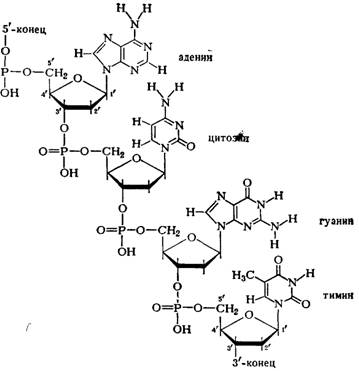
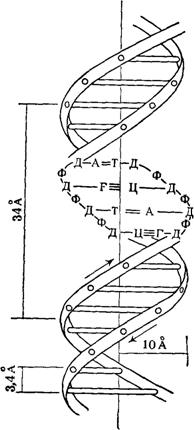
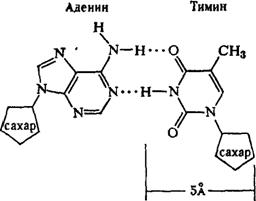
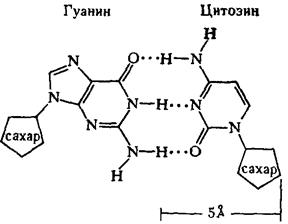
... остатки, соединяющиеся водородными связями с комплементарными основаниями матричной цепи. Рост синтезирующейся цепи происходит в направлении 3'-ОН — 3'-ОН-концам, антипараллельно матричной цепи. Репликация ДНК приводит к удвоению количества генетического материала клетки и, как правило,— к клеточному делению. Поэтому репликация происходит тем чаще, чем короче время генерации вируса или бактерии и ...
0 комментариев