Навигация
1.2 Селекция
Цель селекции – предоставление определенных преимуществ клеткам, необходимым экспериментатору. Существуют четыре группы методов селекции.
1.2.1 Биохимическая селекция, основанная на эндогенных генах
Наиболее широко распространенный пример такого подхода – НАТ-селекция. В присутствии аминоптерина ингибируется синтез de novo предшественников ДНК. Клетки, лишенные фермента тимидинкиназы, не могут утилизировать экзогенный тимидин и гибнут в присутствии аминоптерина. Аналогично, клетки, лишенные гипоксантин-фосфорибозилтрансферазы, не могут усваивать гипоксантин и также нежизнеспособны в присутствии аминоптерина. Соматические гибриды, полученные при слиянии ТК-, НРКТ+-кле-ток с клетками ТК+, HPRT-, могут оказаться одновременно ТК+, HPRT+. Гибридные клетки с таким генотипом способны расти в присутствии аминоптерина, если гипоксантин и тимидин добавлены в среду. Распространен вариант НАТ-метода, когда один партнер по гибридизации не может размножаться in vitro, а другой – является дефицитным по ТК или HPRT. Для исследований в области генетики соматических клеток разработано большое количество систем комплементации. Многие из них основаны на использовании различных ауксотрофных и прототрофных клеточных линий хомячков. Этот подход требует создания подходящих мутантных клеток-реципиентов.
Учитывая важность НАТ-селекции, мы предлагаем вниманию читателей руководство по приготовлению соответствующей среды.
1.2.2 Биохимическая селекция, основанная на экзогенных генах
Этот подход избавляет нас от необходимости выделять специфический мутант, поскольку он подразумевает использование бактериальных генов, которые дают селективные преимущества при их экспрессии в клетках млекопитающих. Для этого конструируют плазмидные и ретровирусные векторы, в которых бактериальные гены сочетаются с промоторами, местами сплайсинга и сигналами полиаденилирования млекопитающих. Введение бактериальных генов в клетки млекопитающих с помощью трансфекции или инфекции приводит к их случайному распределению в геноме реципиента. В качестве примера бактериальных генов, способных обеспечивать селективные преимущества клеток млекопитающих, можно назвать ген Е. coli gpt и генлео. Основной недостаток этого метода – случайное распределение сайтов интеграции; однако последние исследования позволяют надеяться, что с помощью гомологичной рекомбинации удастся осуществлять направленную интеграцию.
1.2.3 Антигены клеточной поверхности
В роли селективного фактора могут выступать антитела. При этом мы получаем возможность выделять клетки с определенным набором антигенов клеточной поверхности. Применение антител лежит в основе ряда методов, среди них отбор клеток с помощью клеточного сортера, розеткообразование с эритроцитами, связанными антителами, избирательное прикрепление клеток к поверхности с иммобилизованными на ней антителами. Хотя эффективность селекции клеток с помощью антител недостаточно высока, тем не менее селекция с использованием антител применяется во всех методах переноса генов, описываемых здесь, за исключением MMGT.
1.2.4 Активированные онкогены
Мутировавшие протоонкогены, особенно члены семейства ras-генов, обеспечивают преимущества в росте клеток млекопитающих и, следовательно, могут быть использованы для селекции.
2. Слияние целых клеток
2.1 Введение
Известно, что при смешивании клеток они могут спонтанно сливаться, однако событие это чрезвычайно редкое. Эффективность слияния может быть существенно увеличена с помощью специфических антигенов. Первоначально для этой цели использовали инактивированный вирус Сендай, однако биологическая неоднородность различных партий инактивированного вируса и громоздкость процедуры слияния, индуцированного вирусом, привели к широкому использованию химического индуктора–полиэтиленгликоля. Метод, представленный ниже, пригоден для слияния клеток как одного, так и разных видов животных. Внутривидовые гибриды формируются с частотой 10~5–Ю-3. Частота слияния клеток разных видов варьирует от 10~7 до 10~5. Клетки, имеющие сходные фенотипы и близкие параметры роста, сливаются с более высокой частотой.
2.2 Получение клеточных гибридов с помощью слияния, индуцированного ПЭГ
Таблица 2. Основные растворы, необходимые для слияния клеток
| Среда для роста клеток Среда для роста клеток без эмбриональной телячьей сыворотки | Обычно мы используем среду Игла, модифицированную Дульбекко, с добавлением эмбриональной телячьей сыворотки, однако состав культуральной среды прямо на процесс слияния не влияет |
| Селективная среда | Культуральная среда, пригодная для селекции гибридных клеток |
| 50%-ный раствор ПЭГ, приготовленный в бессывороточной среде | 5,5 г ПЭГ 4000 н 5 мл бессывороточной среды смешивают и авто-клавируют. Конечный рН доводят до 8,2, раствор нагревают до 37 °С непосредственно перед использованием |
2.3 Возможные ошибки и варианты методики
Некоторые линии клеток сливаются с трудом, если же все-таки необходимы именно такие комбинации, попытайтесь сделать следующее.
1. Варьируйте соотношение родительских клеток в диапазоне 1:10–10:1.
2. Попробуйте разный ПЭГ:
а) поменяйте партию ПЭГ. В наших экспериментах та партия ПЭГ, которая давала хорошие результаты при слиянии одного типа клеток, эффективно работала и с другими. Хорошо зарекомендовавший себя препарат необходимо отделить и аккуратно хранить. По некоторым данным свойства ПЭГ могут быть улучшены использованием растворов, не содержащих солей кальция;
б) варьируйте молекулярный вес. Как правило, повышение молекулярного веса влечет за собой увеличение способности индуцировать слияние. Однако раствор ПЭГ с большим молекулярным весом обладает большей вязкостью, что затрудняет отмывку клеток. Так как ПЭГ является
Для каждой конкретной клеточной линии необходимо подобрать оптимальную концентрацию колцемида и время экспозиции. Образование мини-клеток легко контролируется под фазово-контрастным микроскопом. В удачных экспериментах около 50% всех клеток образуют мини-клетки. Эта частота увеличивается после обработки цитохала-зином В.
Второй день
3. Спустя 16 ч замените среду на среду, содержащую цитоха-лазин В в концентрации 2 мкг/мл; инкубируйте в течение ночи.
4. Исходные градиентные растворы поместите в сосуды с неплотно завинченными крышками и инкубируйте в атмосфере 5% СОг при 37 °С в течение ночи.
5. Заранее нагрейте ротор SW41.
Третий день
6. Приготовление ступенчатого градиента фиколла. Тщательно промойте необходимое количество центрифужных пробирок для ротора SW41 абсолютным спиртом, высушите в перевернутом положении в ламинарном боксе. Приготовьте градиент фиколла, используя уравновешенные исходные растворы.
7. Соберите обработанные колцемидом и цитохалазином клетки и осадите их центрифугированием. Ресуспендируйте в 3 мл 10%-ного фиколла и мягко наслоите на градиент. Заполните центрифужные пробирки раствором без фиколла.
8. Поместите пробирки, содержащие градиент, в нагретый ротор SW41 и поставьте ротор в заранее нагретую до 37 °С ультрацентрифугу.
9. Центрифугируйте 1 ч при 25000 об/мин при 37 °С; используйте минимальное ускорение при разгоне и минимальное торможение, чтобы предотвратить разрушение градиента.
10. Выньте пробирки из центрифуги. Мини-клетки образуют рыхлые полоски в градиенте между 15:16% и 16:17% фиколлом. Осторожно отберите эти полоски, используя стерильную пастеровскую пипетку, вводя ее через верх градиента; поместите мини-клетки в новую, стерильную центрифужную пробирку от ротора SW41 и заполните ее средой для роста клеток. Загрязнение фрагментами цитоплазмы, ядрами и целыми клетками можно контролировать под фазово-контрастным микроскопом.
11. Центрифугируйте при 20 000 об/мин при комнатной температуре в роторе SW41 10 мин при максимальном ускорении и торможении. Эта процедура осаждает мини-клетки и отделяет их от фиколла.
12. Для дальнейшей очистки мини-клеток мы предлагаем три различных приема. Очистка не нужна, лишь когда в распоряжении исследователей имеется четкая система селекции. В этом случае они могут сразу использовать мини-клетки для слияния с клетками-реципиентами.
13. Слияние мини-клеток с целыми клетками. Соберите клетки-реципиенты и отмойте их бессывороточной средой. Добавьте приблизительно 107 клеток-реципиентов к осадку мини-клеток в 2 мл бессывороточной ростовой среды, содержащей 100 мкг/мл фитогемагглютинина. Поместите в пластиковый сосуд с коническим дном и инкубируйте 10 мин при 37 °С.
14. Осадите центрифугированием.
15. Процедуру слияния мини-клеток с целыми клетками проводите с помощью ПЭГ, как указано в соответствующей методике.
3. Перенос генов, опосредованный хромосомами JCMGTJ
3.1 Введение
Метод CMGT может быть использован для переноса фрагментов хромосом из ядер клеток одного типа в ядра клеток другого типа. Теоретически клетки любого типа могут быть использованы как в качестве доноров, так и в качестве реципиентов хромосом. Однако на практике возможность применения метода определяется наличием подходящих реципиентных линий, обладающих повышенной способностью акцептировать чужеродную ДНК.
Высокая частота трансфекции может быть достигнута при использовании в качестве реципиентов иммортализованных мышиных клеток. Клеткам хомячка и иммортализованным человеческим клеткам обычно присуща более низкая частота трансфекции. Правда, недавно полученные результаты свидетельствуют о том, что человеческие клетки линии EJ способны трансфицироваться с высокой частотой. В роли донора с одинаковым успехом могут выступать самые разнообразные клеточные линии – как суспензионные, так и субстрат-зависимые. Предпочтительнее использовать в качестве донорных те линии, клетки которых легче культивировать и получать в больших количествах. Ниже описываются общие процедуры, обеспечивающие выделение донорных хромосом и перенос фрагментов этих хромосом в реципиентные клетки путем CMGT.
Похожие работы
... : генный, хромосомный и геномный. На каждом из них проявляются основные свойства материала наследственности и изменчивости и определенные закономерности его передачи и функционирования. 4. Генный уровень организации генетического аппарата Элементарной функциональной единицей генетического аппарата, определяющей возможность развития отдельного признака клетки или организма данного вида, ...
... , доступные генетическому анализу — это прежде всего скорость и полнота лизиса инфицированных клеток и круг бактерий-хозяев, поражаемых фагами. Широкое распространение в генетическом анализе бактериофагов получили мутанты с условным проявлением. Это мутанты, чувствительные к повышению и понижению температуры, — так называемые термочувствительные (ts) и холодочувствительные (cs). Они нормально ...
... 1972 год, когда в Стенфордском университете П. Берг и С. Коэн с сотрудниками создали первую рекомбинантную ДНК, содержавшую фрагменты ДНК вируса SV40, бактериофага и E. coli. Генетическая инженерия Одним из разделов молекулярной генетики и молекулярной биологии, который нашел наибольшее практическое приложение, является генная инженерия. Генная инженерия – это сумма методов, позволяющих ...
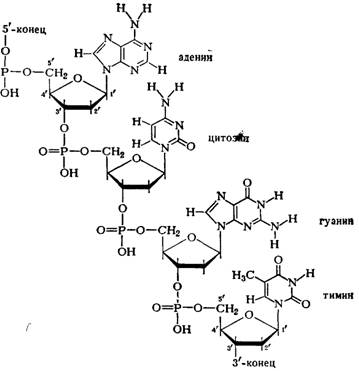
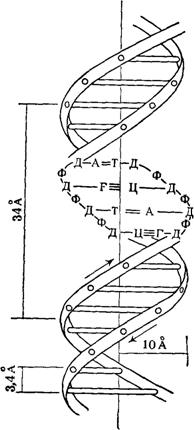
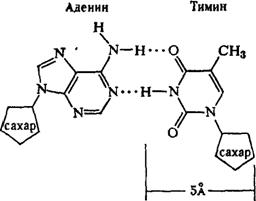
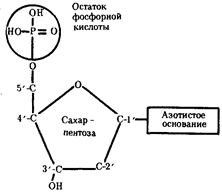
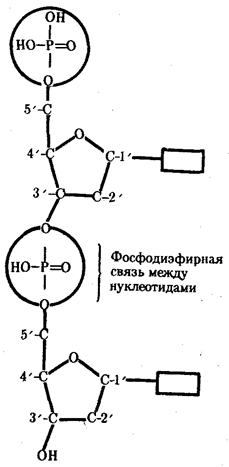
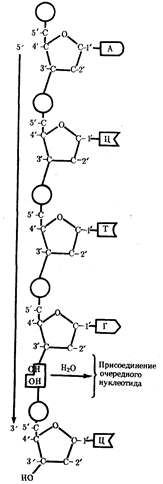
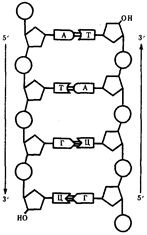
... остатки, соединяющиеся водородными связями с комплементарными основаниями матричной цепи. Рост синтезирующейся цепи происходит в направлении 3'-ОН — 3'-ОН-концам, антипараллельно матричной цепи. Репликация ДНК приводит к удвоению количества генетического материала клетки и, как правило,— к клеточному делению. Поэтому репликация происходит тем чаще, чем короче время генерации вируса или бактерии и ...
0 комментариев