Навигация
Рекомбинация и генетический анализ бактериофагов
23421
знак
0
таблиц
0
изображений
Министерство сельского хозяйства РФ
ФГОУ ВПО
Оренбургский государственный аграрный университет
Кафедра микробиологии
Реферат
по генетике микроорганизмов на тему:
Рекомбинация и генетический анализ у бактериофагов
Выполнил: студент ΙV курса
ФВМ и Б специальности
«Микробиология» Акжигитов А.С.
Проверил: преподаватель кафедры
микробиологии Капустина О.А.
Оренбург 2010
Содержание
Введение
1. Рекомбинация у бактериофагов
2. Генетический анализ бактериофагов
Заключение
Список литературы
Введение
В основе наследственной изменчивости бактериофагов помимо мутации лежит рекомбинация. Рекомбинация у бактериофагов – это физическое взаимодействие геномов в смешанно-инфицированных клетках. При этом происходит обмен генетическим материалом или его частью между двумя (часто близкими) отличающимися по наследственным свойствам вирусами. Рекомбинация между родительскими геномами приводит к возникновению новых сочетаний генов в дочерних геномах, т. е. формированию нового генома. Рекомбинация путем включения одной молекулы ДНК в другую происходит между вирусной ДНК и ДНК клетки-хозяина. На этом основан метод искусственной рекомбинации молекул ДНК
В последнее десятилетие ДНК-вирусы человека и животных стали привлекать внимание генетиков как модель для изучения рекомбинации в клетках животных, особенно в связи с деталями механизма разрыв – воссоединение. В силу известной гетерогенности вирусных популяций в процессе их размножения в клетках могут создаваться условия не только для генетических, но и негенетических взаимодействий. Последние вызывают большое разнообразие фенотипических изменений, которое, маскируя истинный генотип вируса, влияет на результаты генетических исследований. Поэтому негенетические взаимодействия вирусов традиционно включаются в раздел анализа изменчивости вирусов в результате их взаимодействия друг с другом. Негенетические взаимодействия описаны у широкого круга вирусов и могут происходить между гетерогенными их группами и даже между РНК- и ДНК-содержащими.
1. Рекомбинация у бактериофагов
При взаимодействии бактериофагов друг с другом наследуемые изменения возникают в основном как результат истинных рекомбинаций. У вирусов животных и человека в зависимости от физической организации генома рекомбинация осуществляется двумя механизмами. Бактериофаги, геном которых представлен линейной молекулой ДНК или РНК, используют механизм интрамолекулярной рекомбинации, известный как механизм «разрыв — воссоединение». Бактериофаги с сегментированным геномом (ортомиксовирусы, реовирусы, буньявирусы, аренавирусы) используют механизм случайной пересортировки сегментов (генов) без разрыва ковалентных связей.
Механизм «разрыв — воссоединение» впервые сформулирован А. Херши в связи с анализом потомства смешанного заражения h-и r-мутантами фага Т2. Обнаружив, что в этом потомстве кроме частиц двух родительских типов присутствуют частицы h+r+ и hr, он предположил, что геном бактериофага состоит из расположенных в линейном порядке генов, каждый из которых несет генетическую информацию о каком-либо признаке вируса. При смешанном заражении бактериальной клетки два «сосуществующих» фаговых генома могут обмениваться между собой гомологичными участками в результате разрыва двух линейных структур в точно соответствующих точках между генами, определяющими г+- и h+-признаки, и последующего перекрестного воссоединения фрагментов. Такой перекрест или кроссинговер генетического материала может происходить в любой точке двух различных генетических структур.
Гипотеза «разрыв — воссоединение» была подтверждена позже на опытах по смешанному заражению меченым фагом. R. Holliday, Н. Potter, D. Dressier предложили универсальную модель одно- и двухцепочечного обмена между двумя линейными молекулами (рис. 1), согласно которой рекомбинация — такой же многоэтапный процесс, как и мутация. Она начинается с разрыва цепей ДНК каждого партнера с помощью нуклеаз, на этом месте происходят перекрещивание цепей и сшивание концов, затем точки скрещивания перемещаются за счет расплетения и сплетения спиралей и вращаются вокруг точки пересечения, создавая при этом симметрическую структуру — «хи»-интермедиат. В зависимости от того, на какую ось интермедиата (вертикальную или горизонтальную) действуют нуклеазы, образуемый после разделения и сшивания рекомбинант будет гетерозиготным либо по одной, либо по двум цепям.
Из ДНК-вирусов с линейной молекулой генома рекомбинационный процесс хорошо изучен герпес- и аденовирусов, которые рекомбинируют как с близкородственными, так и неродственными вирусами.
Генетическая рекомбинация между аденовирусами человека происходит с высокой частотой при продуктивной инфекции культур клеток. Причем рекомбинация наблюдается внутри одного серотипа или между близкородственными серотипами одной подгруппы, что связывают с отсутствием гомологичных последовательностей в геномах вирусов разных подгрупп, хотя общая их организация сходна. Было опрделено, что на рестрикционной карте ts+-peкомбинантов Ad5 и Ad2 предполагаемые сайты находятся в фрагменте размером около 20 п. н., занимающем участок соединения С-конца гена PV1 и N-конца гексонового гена. Сравнительное секвенирование аналогичных фрагментов трех независимых скрещиваний выявило, что в каждом случае непосредственное участие в рекомбинационном процессе принимала лишь небольшая его часть (от 45 до 156 п. н. в длину), соответствующая участкам полной гомологии ДНК.
Герпесвирусы занимают более выгодное положение: ДНК разных серотипов способны в одинаковой степени трансфицировать культуры клеток, имеют значительные участки гомологии и рекомбинируют с большой частотой. Создавались как межтиповые, так и внутритиповые рекомбинанты, с помощью которых картировались генные функции, контролирующие репликацию вируса, морфологию бляшек, резистентность к антивирусным лекарствам. R. Thompson котрансфекцией ДНК одного штамма с разными рестрикционными фрагментами другого получили рекомбинант, позволивший локализовать функцию повышения нейровирулентности.
Метод получения межтиповых рекомбинантов полиовирусов разработан V. Agol с сотрудниками на основе коинфицирования клеток gs-мутантом одного серотипа и gr-мутантом другого с последующей селекцией gr-клона из урожая двойной инфекции. Таким способом получена серия межтиповых рекомбинантов, сайты скрещивания которых были локализованы в центральной части генома между локусом антигенной специфичности (5'-сторона) и чувствительностью к гуанидину (3'-сторона). Определены первичные структуры скрещиваемых регионов, длина которых варьирует между 2 и 32 нуклеотидами. Сайты скрещивания оказались распределенными по геному неравномерно. Так, внутри гена полипептида 2А обнаружен только один такой участок, в то время как в других регионах выявлялись явные их скопления, что указывало на существование предпочтительных сайтов для рекомбинации.
Используя тот же принцип, V. Agol е. а. (1984) получали внутритиповые рекомбинанты между аттенуированным и нейровирулентным штаммами полиовирусов, изменения нейровирулентности которых определили интрацеребральным заражением обезьян. Показано, что рекомбинанты, унаследовавшие 5'-половину генома от вирулентного родителя, проявили нейровирулентный фенотип независимо от происхождения 3'-половины, а рекомбинанты с 5'-геномной половины от аттенуированного родителя имели аттенуированный фенотип. Следовательно, большие детерминанты нейровирулентности находятся на 5'-половине генома, а на 3'-половине — минорные или модулирующие детерминанты.
Таким образом, детальный анализ межтиповых и внутритиповых рекомбинантов полиовирусов представил окончательные доказательства истинности рекомбинации, опроверг случайность процесса, показал, что скрещивания происходят в определенных, хотя и многих сайтах.
Интромолекулярная рекомбинация зафиксирована и у вируса гриппа. Были обнаружены последовательности сегментов ДИ РНК, составленные из последовательностей от двух нормальных геномных сегментов.
Кроме того, К. Shimizu е. а. представили доказательства существования внутрисегментной комплементации между ts-мутантами вируса гриппа. По их данным, 83 ts-мутанта образовывали 13 комплементационных групп и 8 рекомбинационных. При этом четыре рекомбинационные группы включали вирусы, представляющие более чем одну комплементационную группу, а группа Н – в каждом комплементационном члене имела по четыре ts-локуса. Мутации с внутрисегментной комплементацией обнаружены в большинстве генов (РЗ, PI, P2, NA, NP и NS).
Относительно механизма внутрисегментной комплементации высказывались различные гипотезы. Считалось, что она проявляется: 1) если сегмент полицистронен (известно, что два сегмента (7, 8) генома вируса гриппа А кодируют по два белка Ml, M2 и NS1, NS2, но, внутрисегментная комплементация отмечена и при мутациях в генах, содержащих информацию для единственных белков) ; 2) если множественный белок состоит из смеси аллельных белковых субъединиц, кодируемых двумя комплементарными родительскими генами; 3) если родительские комплементарные вирусы несут мутации в генах, кодирующих белки с несколькими функциональными доминантами.
Много неясного остается и в другом механизме рекомбинации вирусов человека и животных — реассортименте. У вирусов с сегментированным геномом каждый сегмент — это независимая и самостоятельная молекула. Полагают, что в зараженных клетках существует активный механизм, регулирующий, чтобы каждый вирион получил из клеточного пула по одной копии всех сегментов. Можно предполагать, что перекомбинация генов осуществляется на стадии морфогенеза вирусов и этот процесс также зависит от клеточных факторов. К тому же выявить участие в этом процессе вирионных белков или каких-либо последовательностей нуклеотидов, способствующих ему, не удалось.
Из вирусов с сегментированным геномом рекомбинационный процесс наиболее детально изучен у ортомиксовирусов и реовирусов. Исследования на ортомиксовирусах начаты F. Burnet (1960) еще задолго до установления сегментированности их генома. Он получил ряд основополагающих данных, вплоть до предсказания структуры генома. В этих опытах потомство смешанной инфекции двумя штаммами, один из которых (WSN) был нейротропным, оказалось способным вызывать смертельную инфекцию при заражение мышей в мозг, но имело серологическую характеристику пневмотропного MEL штамма. При прямой и обратной рекомбинации этих штаммов, различающихся по семи признакам, обмен происходит как бы по двум сцепленным признакам. Одну группу сцепления составляли признаки, определяющие серологическую специфичность, термостабильность гемагглютининов, отношение к ингибирующему действию муцина овцы и патогенность для мышей при интраназальном заражении. Другая группа включала отношение к ингибирующему действию овомуцина, патогенность для куриных эмбрионов и нейропатогенность для мышей.
G. Hirst высказывал предположение, что вирусы гриппа могут скрещиваться по двум механизмам: 1) обмен полноценными сегментами с высокой частотой рекомбинации; 2) кроссинговер гомологичными сегментами с низкой частотой рекомбинации. Теперь имеется множество прямых доказательств способности вируса гриппа использовать при рекомбинации как интрамолекулярный механизм, так и перекомбинирование генов. Они основаны на сравнительном изучении электрофоретической подвижности сегментов генома и его продуктов, а также на их секвенировании.
Впервые антигенные гибриды из вируса разного серотипа выделены в лабораториях М. И. Соколова и Е. Kilbourne. Все рекомбинанты оказались асимметрическими в «антигенной реактивности», т. е. проявляли большую антигенность, унаследованную от одного из родительских штаммов. Независимо от этих исследований В. Tumova, N. Pereira представили доказательства рекомбинации между вирусами гриппа человека А2 и птиц АО. Двойная антигенность потомства устанавливалась в вирусспецифической реакции связывания комплемента. Р. Вебстер получил антигенные гибриды при смешанном заражении куриных эмбрионов различными штаммами вирусов гриппа А млекопитающих и птиц. Поверхностные антигены этих вирусов состояли из нейраминидазы одного родительского штамма и гемагглютинина другого.
Лабораторные доказательства антигенной гибридизации среди разных подтипов вирусов гриппа А послужили основой гипотетических механизмов, объясняющих антигенную лабильность и потенциал чрезвычайной изменчивости этого агента.
Рекомбинация неповрежденных участков генетического материала лежит в основе и другого генетического взаимодействия — множественной реактивации, заключающейся в формировании полноценного вируса за счет взаимного обмена активными участками генома. Таким образом, если кроссреактивация включает «ремонт» нарушений или неадекватности генома генетическим вкладом от инфекционного вируса, то множественная реактивация подразумевает взаимную помощь или «кооперацию» двух пораженных геномов без участия инфекционного вируса. Эти типы реактивации показаны в разных вирусных системах и наблюдаются между близкородственными вирусами и разными штаммами одного и того же вируса, между вирусами разных серотипов и неродственными вирусами, например Adl2 и ОВ-40. Примером межтиповой множественной рекомбинации служит результат опыта с облученными реовирусами млекопитающих трех серотипов. В основе феномена, скорее, лежит обмен неповрежденными сегментами генома, а не внутримолекулярная рекомбинация, причем перераспределение сегментов геномов разных серотипов может быть источником антигенных вариантов. А ts-мутанты З серотипа реовируса млекопитающих и птичьи реовирусы не взаимодействовали в клетках L, что указывает на их генетические различия.
Рекомбинантные механизмы лежат в основе возникновения и гетерозигот. В классической генетике этим термином определяют организм, в диплоидном наборе хромосом которого содержатся два различных аллеля какого-либо гена. У большинства вирусов животных такого рода гетерозиготность не наблюдается, поскольку их геномы гаплоидны. Гетерозиготными по всем маркерам могут быть ретровирусы, содержащие две копии геномной РНК. Гетерозиготность ретровирусов связывают с рекомбинационным процессом. У некоторых ДНК-вирусов (например, герпеса), имеются повторяющиеся последовательности и эти вирусы частично диплоидны. В этих диплоидных локусах также может возникать гетерозиготность. Так, выделено несколько типов рекомбинантов при скрещивании вирусов герпеса типа 1 и 2, которые содержали гетерозиготные терминальные повторяющиеся последовательности.
Похожие работы
... , поскольку во время репликации в системе хозяин-вектор иногда происходит клонирование случайных последовательностей, образовавшихся в результате рекомбинаций, делеций или мутаций. Основной подход состоит в использовании клонированной ДНК в качестве зонда для анализа продуктов эндонуклеазного расщепления суммарной геномной ДНК, из которой произошел данный клонированный сегмент. Такой подход ...
... методы составления генетических карт. 1. Перенос генетического материала у актиномицетов 1.1 Перенос генетического материала с помощью плазмид Это наиболее часто встречающийся способ переноса генетического материала у актиномицетов. Линейные плазмиды актиномицетов были обнаружены раньше, чем линейные хромосомы. Впервые они были описаны у одного из видов актиномицетов в конце 70-х – начале ...
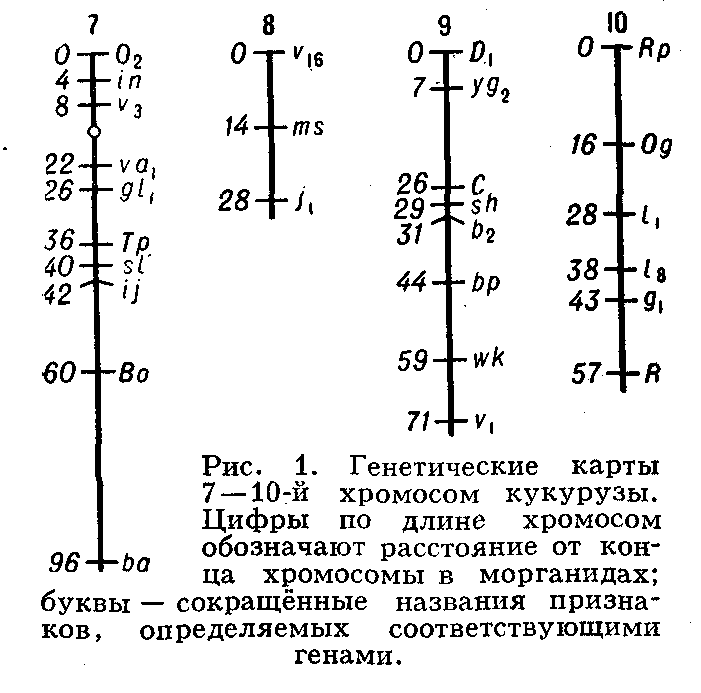
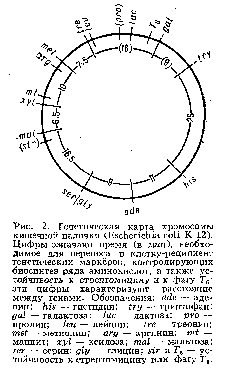
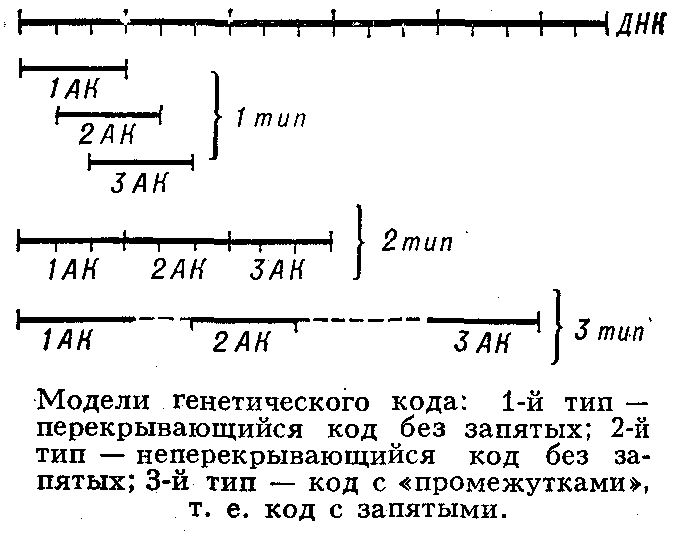
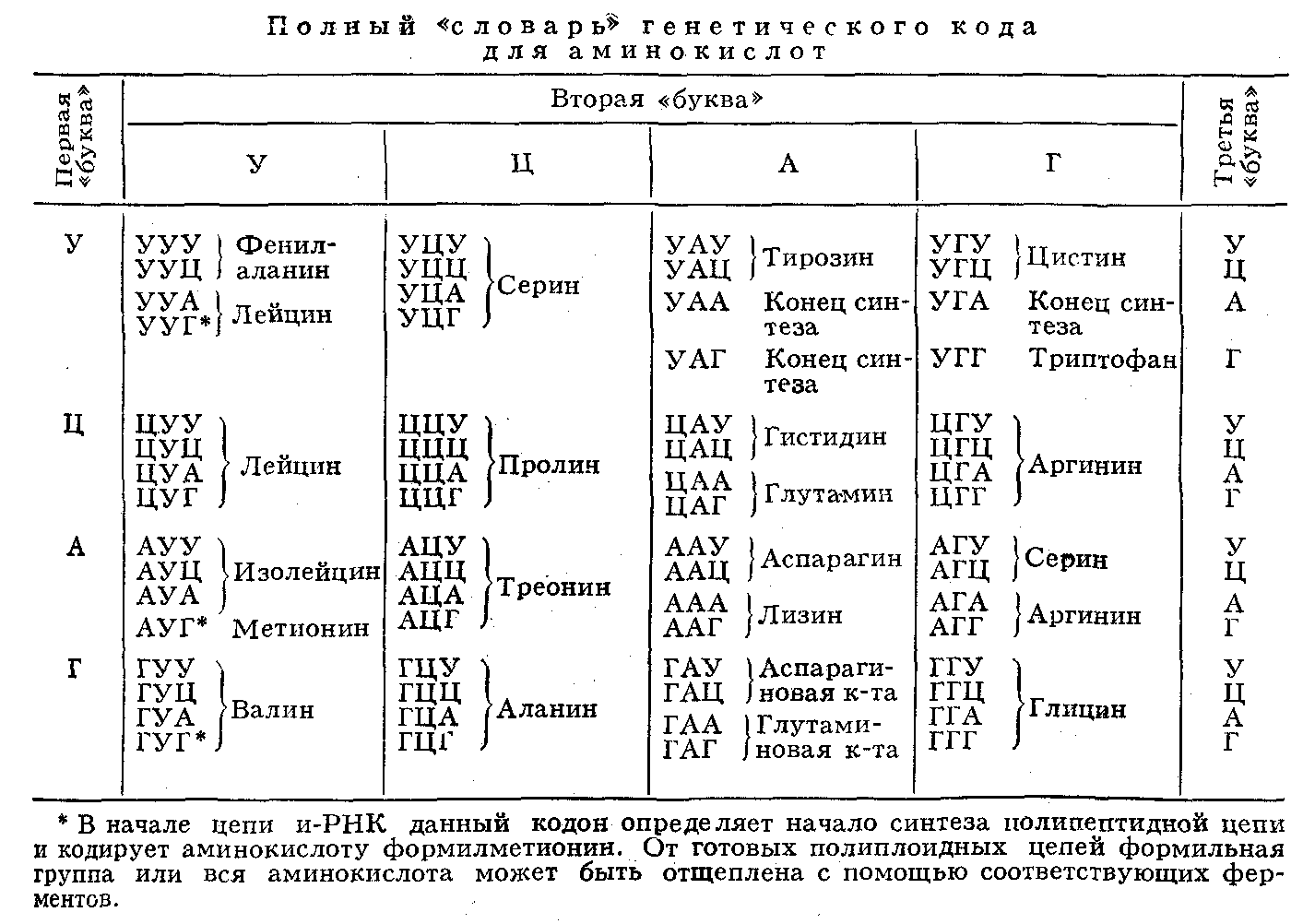
... живые организмы-и удивительное многообразие генов, кодирующих эти белки. В геноме каждого человека есть какие-то области, определяющие его индивидуальность. Некоторые гены человека отличаются от генов крысы всего несколько нуклеотидами-знаками генетического кода. Другие гены у них разные, но одинаковые у двух людей. Изменчивость, связанная с существованием генов , подобных генам группы крови у ...
... один или несколько генов в чистом виде. Такое относительно простое применение принципов и методов, разработанных Важные открытия Как это часто бывает при развитии науки, методологии рекомбинантных ДНК прокладывали путь многие открытия, казавшиеся ранее незначительными и не имеющими отношения к делу. Одним из таких открытий была трансдукция. Другие связаны с выделением и изучением свойств ...
0 комментариев