Навигация
Презентация микобактериальных липидов группой I CD1 молекул
70973
знака
0
таблиц
4
изображения
4.2 Презентация микобактериальных липидов группой I CD1 молекул
Исследования, проведенные Порцелли, Бреннер, Кроненберг и Модлин доказали, что CD1a, CD1b, CD1c молекулы презентуют микобактериальные гликолипиды клеточной стенки. Микобактерии обладают клеточной стенкой, богатой гликолипидами, что очень важно для их устойчивости. Было доказано, что CD1 молекулы первой группы презентуют на Тαβ-клетках гликолипиды фосфоиннозитолманнозиды (PIM), липоарабиноманнан, (LAM) миколовые кислоты и гексозо-1-фосфоизопреноиды. Не было идентифицировано ни одного бактериального антигена, презентуемого CD1 молекулами второй группы.
CD1d-рестриктированные клетки имеют уникальный фенотип. Они экспрессируют на поверхности CD3 комплекс также хорошо, как и маркеры для NK, NK1-клеток. Более того, они экспрессируют высоко специфичный ТКР репертуар, включающий Vα14Jα281 комбинацию у мыши и гомологичную Vα24JαQ у человека. Поэтому считается, что NK Т-клетки похожи. Этот факт также подтверждается тем, что они реагируют с αGalCer.
4.3 Внутриклеточная локализация CD1 молекул
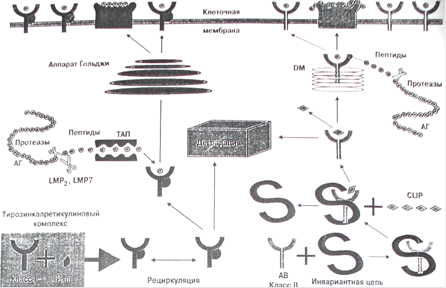
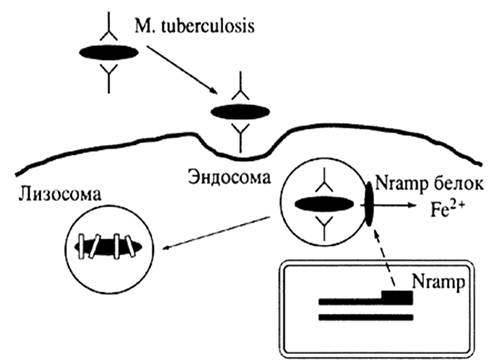
Фагоцитированный макрофагами бактериальный патоген попадает в фагосому, которая затем проходит через несколько стадий. Главные шаги, которые проходит фагосома:
1.Ранняя эндосомальная стадия, на которой фагосома сближается и трансферрином.
2.Поздняя эндосомальная стадия, на которой фагосома закисляется до оптимальной для лизосомальных ферментов pH.
3.Фагосома, в которой идет внутриклеточноке "пищеварение".
Микобактерии мешают созреванию фагосом, хотя механизм этого пока еще не известен. Микобактериальные фагосомы не до конца закислены, но бактерии получают доступ к трансферрину, который является главным депо железа не только для клеток хозяина, но и для микобактерий. Трансферрин и его рецептор двигаются вдоль ранней переработанной эндосомы, отдает готовое железо и затем возвращается на клеточную поверхность. Показано, что микобактериальные гликолипиды в самом деле нагружаются на CD1 молекулы в течении микобактериальной инфекции, эти молекулы могут быть найдены в компартментах. Внутриклеточная локализация CD1 молекул и микобактерий, покрывающих их была анализирована с помощью конфокусной сканирующей лазерной микроскопии дендритных клеток зараженного человека. При использовании различных внутриклеточных меркеров было найдено, что три CD1 молекулы первой группы локализованы в клетки по-разному. CD1а молекула по распределению похожа на МНС первого класса и точно также экспрессирована на клеточной поверхности. Кроме того, CD1а молекула была локализована на ранних рециклирующих эндосомах положительных для GT-Pase ARF6 и для гранул Бирбека клеток Лангерганса. CD1b и CD1c молекулы были найдены в поздних эндосомально-лизосомальных везикулах. Данные компартменты также служат как "доки" для разгрузки пептидов в МНС молекулы второго класса. В противопоставление CD1b, CD1с молекула обильно презентуется на плазматической мембране и в Tf-лабильных ранних эндосомах. Кроме того, было найдено, что многочисленные гликолипиды, включая LAM и PIM, отщепляются от микобактерий внутри фагосом и транспортируются из фагосомы в поздние эндосомы и лизосомы. CD1b и CD1с молекулы были также обнаружены в этих компартментах. CD1с молекулы могут также взаимодействовать с микобактериальными гликолипидами, потому что в зараженных клетках они были обнаружены в микобактериальных фагосомах, задержанных на ранней стадии активации [24].
CD1b молекулы были первоначально найдены в зрелых фаголизосомах. Как ранее было показано, созревание фагосом сопровождается потерей жизнеспособности микобактериями, поэтому эти фаголизосомы скорее всего состоят из нежизнеспособных микобактерий. Дальнейшие эксперименты показали, что зараженные клетки хозяина отщепляют гликолипиды, которые могут быть поглощены дендритными клетками, лежащими в их окрестности. Данные открытия объясняют транспорт антигенных гликолипидов из макрофагов в ДК, или из главных хозяйских клеток к главным АПК, экспрессирующим CD1 молекулы, в эксперименте in vitro. Предварительные данные свидетельствуют, что внутриклеточные везикулы различных размеров, такие как экзосомы и апоптические пузырьки, принимают участие в данном транспорте. Во время микобактериальной инфекции экспрессия на поверхности молекул МНС первого и второго класса и CD1b нерегулируема.
4.4 Рецептор для транспорта АГ
Маннозо-мембраные рецепторы (PPR) играют роль в понимании (распознавании) и презентации очищенного LAM с помощью CD1b. Этот R был идентифицирован в ранних эндосомах, но не в микобактериальных фагосомах, что доказывает его участие в распознавании чистых гликолипидов. РРR CD14 связывает не только липополисахариды грам "-" бактерий, но также некоторые микобактериальные гликолипиды. Этот R путешествует через фагосомы и поздние эндосомы-лизосомы зараженных микобактериями макрофагов. Отсюда следует, что CD14 может принимать участие в транспорте гликолипидов из фагосом в другие внутриклеточные компартменты. Так как CD14 экспрессируется только на макрофагах, то дендритные клетки, несущие CD1 молекулу, должны использовать другие R, для транспорта гликолипида. Предполагается, что гликолипиды транспортируются из зараженных макрофагов в незараженные ДК для презентации, а также, что CD14 может принимать участие в транслокациях гликолипида внутри макрофага. Как было показано для LAM, гликолипиды могут встраиваться в мембрану хозяйской клетки и мигрировать вдоль фосфолипидного бислоя. В фагосомах CD1 может принимать гликолипиды из депо. Здесь загрузка CD1а и CD1с (но не CD1b) независима от низкого рH. Расщепленные микобактериальные гликолипиды могут быстро связываться с экспрессированными на клеточной поверхности молекулами CD1 с помощью механизма экстрацеллюлярной загрузки – этот процесс постулирован для CD1а и, возможно, CD1с.
Считается, что загружающие гликолипиды в CD1 молекулы могут включать шаперон-подобные молекулы для облегчения связывания половины гидрофобного липида с гидрофильной антиген-связывающей щелью. Также исследуется, какие микобактериальные гликолипиды нуждаются в переработке до того, как они будут связаны и презентованы молекулами CD1. было показано, что презентация гликолипида молекулами CD1b (но не CD1а) включает в себя транспорт гликолипидов в лизосомальные компартменты. Более того, эндосомально путешествующие CD1b, CD1с и CD1в молекулы содержат YXXZ эндосомально-целевую последовательность (Z содержит большую гидрофобную часть цепочки). Этот целевой мотив помогает ассоциации CD1 молекул с адапторным протеином (АР) их корректной внутриклеточной сортировке. При мутации данной последовательности отменяется презентация антигена молекулой CD1b. Кислая среда внутри лизосом может облегчить расщепление антиген – связывающий щели CD1 и обрезанию гликолипидов лизосомальными гликозидазами и липазами. Действительно, при проведении экспериментов с меченными радиоактивными атомами микобактериями было доказано, что микобактериальные гликолипиды ферментативно изменяются на их пути из фагосом.
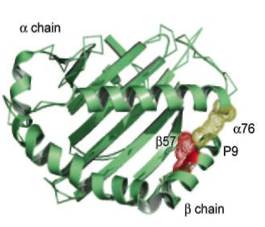
Антиген-связывающая щель CD1 молекул, возможно, связывает две гидрофобные цепочки жирных кислот гликолипидов, тогда как гидрофильная углеводная часть высовывается для распознавания Т-клетками (рис. 4). Ферментативная модификация углеводной части может привести к дифференциации Т-клеточных эпитопов и, следовательно, к антигенной специфичности, несмотря на то, что CD1 молекулы неполиморфны. Ферментативная модификация жирных кислот может улучшить аккомодацию гликолипидов в щели CD1 молекул [23].
Похожие работы

... их гаплотических сочетаний у больных туберкулезом представителей русской этнической группы Челябинской области При сравнительном анализе особенностей распределения специфичностей генов HLA II класса у больных туберкулезом и здоровых представителей русской этнической группы Челябинской области были установлены следующие факты: у больных туберкулезом и здоровых пациентов были выявлены различия в ...
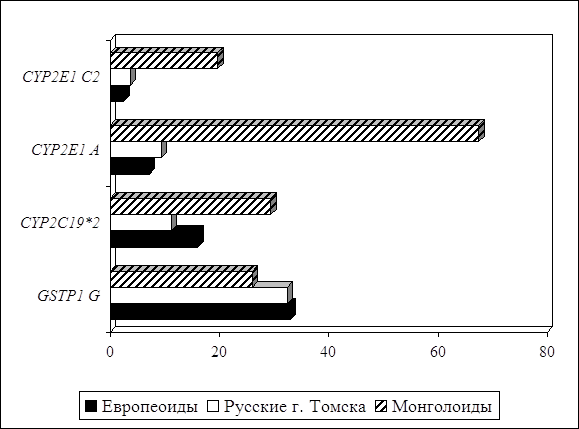
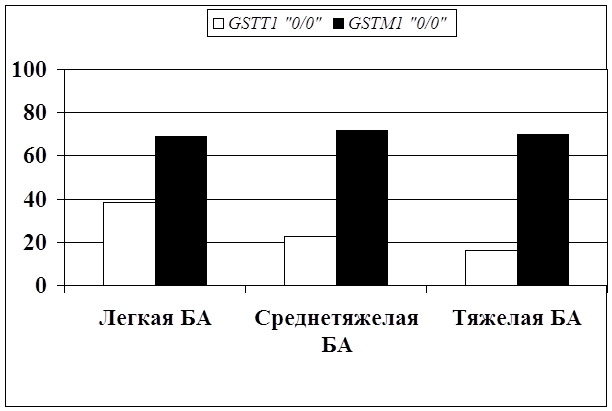
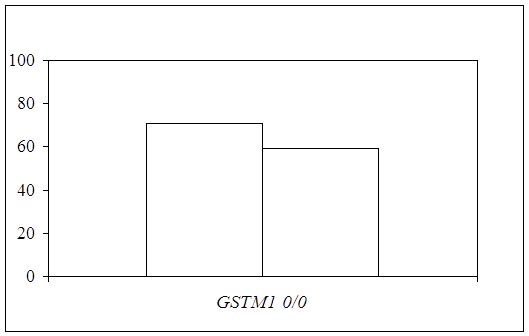
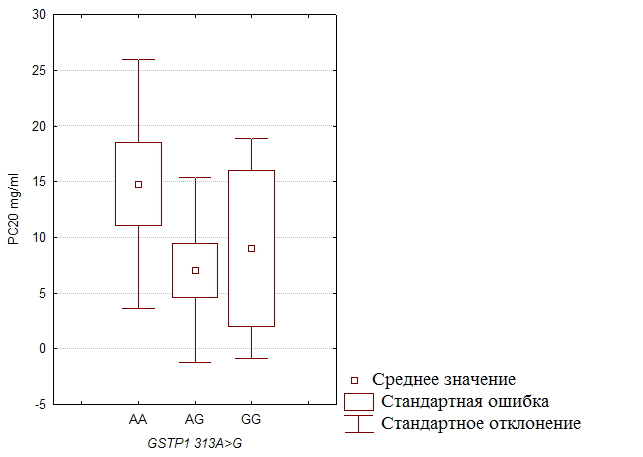
... препаратов. Установлена связь полиморфизма 313A>G гена GSTP1 с изменчивостью уровня аланинаминотрансферазы (р=0,021). 7. Выявлены различия в структуре генетической подверженности к бронхиальной астме и туберкулезу по генам ферментативной системы метаболизма ксенобиотиков: гены GSTM1, CYP2E1 и CYP2C19 связаны с бронхиальной астмой и значимыми для заболевания качественными и ...
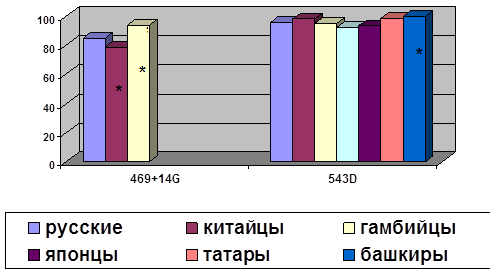
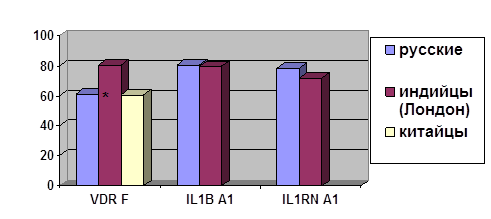
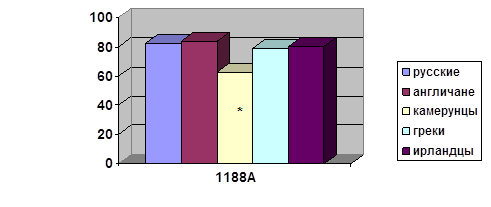
... о связи между частотой аллелей генов-кандидатов ТБ и особенностями распространения этого инфекционного заболевания. Выводы Выявлены особенности в распределении частот аллелей, изученных генов–кандидатов подвержености к туберкулезу, у русских жителей г. Томска по сравнению с другими популяциями мира. При сравнении с тувинцами показаны отличия в распределении генотипов и частот аллелей по всем ...
... фиброза и деформации лёгочной ткани. Фиброзно-кавернозный туберкулёз значительно хуже подвергается заживлению, чем кавернозный. Цирротический туберкулёз. Цирротический туберкулез лёгких характеризуется развитием в лёгочной ткани грубого, деформирующего орган склероза (цирроза), бронхоэктатических, посткавернозных типа кист полостей, эмзифематонных булл или каверн без признаков прогрессирования. ...
0 комментариев