Навигация
Иммунный ответ на персистирующие микобактерии
70973
знака
0
таблиц
4
изображения
3.4 Иммунный ответ на персистирующие микобактерии
Пока мало известно об иммунных механизмах, которые вовлечены в ответ против персистирующих микобактерий. В последних работах с использованием методов молекулярных отпечатков было показано повторное заражение иммуннокомпетентных людей новыми штаммами M. Tuberculosis. Это доказывает, что иммунитет к туберкулезу может быть неполным и что повторное заражение играет немаловажную роль.
Роль адаптивного иммунного ответа при персистенции микобактерий очень важна. Есть некоторые свидетельства, что CD8+ Т-клетки секретируют большую часть ИФНγ. Во время острой фазы болезни большую часть ИФНγ продуцирует CD4+ Т-клетки, доказывая различную активацию Т-лимфоцитов в эти фазы инфекционного процесса. В дополнение к продукции ИФНγ активные CD8+ Т-клетки контролируют персистентные инфекции.
Продукция иммунносупрессорных цитокинов, таких ИЛ10 и ТФРβ, была описана у людей с активным туберкулезом и повышение продукции ИЛ10 в легких у мышей с хронической микобактериальной инфекцией. Это свидетельствует о том, что ИЛ10 может способствовать реактивации хронического туберкулеза.
Глава 4. CD-1 и CD-1-рестриктированные Т-клетки при инфекциях, вызванных внутриклеточными бактериями
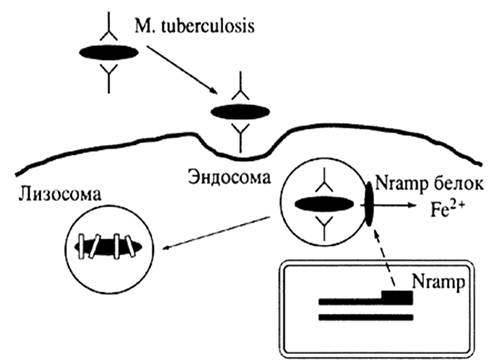
Гликолипид-специфичные, CD-1a-, b-, c- зависимые цитотоксические Т-клетки вовлечены в ответ организма против туберкулеза (рис. 3). Молекулы CD-1 презентуют микобактериальные гликолипиды из различных внутриклеточных участков зараженной клетки. Во время микробной инфекции CD-1d-зависимые натуральные киллеры быстро продуцируют цитокины и выполняют регуляторную функцию [23].
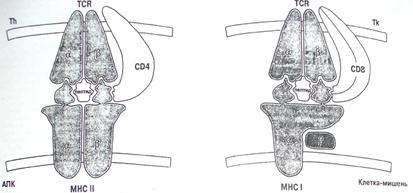
Участие главного комплекса гистосовместимости (МНС) в Т-клеточном ответе было показано на вирусных системах, но его важность в бактериальных инфекциях и в ответе против простейших бесспорно. МНС-II-рестрикрированные CD8+ Т-клетки играют важную роль в защите организма против бактерий и простейших, тогда как МНС-I-рестриктированные CD8+ Т-клетки важны в случаях вирусных инфекций. Первые Т-клеточные популяции выполняют хэлперную функцию, выделяются цитокины для активации макрофагов и/или В-клеток, тогда как последующие, главным образом, действуют, убивая зараженные клетки хозяина. Во время некоторых бактериальных инфекций вместе с обычными CD4+ и CD8+ Т-клетками также есть необычные Т-клетки для содействия в достижении устойчивости: это Тγδ-лимфоциты, которые распознают фосфолипиды и Тαβ-лимфоциты, которые контролируются неклассическими МНС и МНС-похожими молекулами. Т-клетки, контролируемые CD-1 молекулами и противодействующие бактериальным гликолипидам, также являются необычными Т-клетками.
4.1 Атигенпрезентующие молекулы
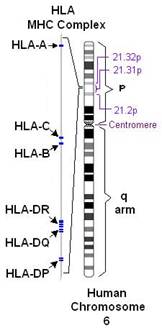
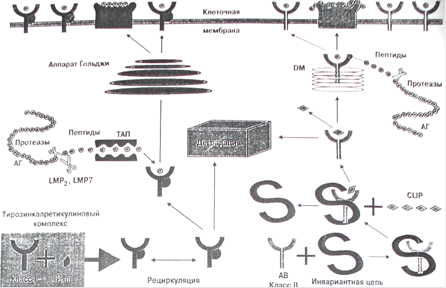
В цитоплазме белки разрушаются протеазами. Образовавшиеся пептиды затем трансформируются в эндоплазматическом ретикулуме специальными транспортными молекулами так называемыми транспортерами переработанного антигена (ТАР). В эндоплазматическом ретикулуме эти пептиды затем обрезаются для упаковки в щель, сформированную первым и вторым доменом тяжелой цепи МНС-I. После ассоциации с β2 микроглобулином (β2m), комплекс МНС-I- β2m транспортируется на клеточную поверхность, где презентует пептиды для CD8+ Т-клеток. Молекула МНС-II состоит из двух тяжелых цепей: α-цепь и β-цепь, первые домены обоих цепей образуют щель, в которую загружается пептид в поздних эндосомально-лизомальных отделах. Пептиды, которые презентуются молекулами МНС-II, образуются в эндосомально-лизомальной системе под действием протеаз (таких как аспарагинилпептидаза, катепсины). Благодаря большому полиморфизму молекул МНС, каждая обладает уникальным гаплотипом. Это гарантирует то, что огромное множество антигенных пептидов будет презентовано и исключает возможность, что какой-то патоген избежит распознавания Т-клетками. Различные антигенные пептиды выбираются разными видами МНС. В противоположность классическим молекулам МНС неклассические МНС-Ib и CD-1 не обладают полиморфизмом. Поэтому антигенные лиганды, презентуемые данными молекулами Т-клеткам, должны быть более устойчивыми [24].
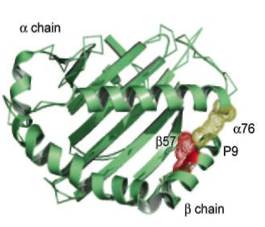
До недавнего времени считалось, что Т-клетки оказывают влияние на пептидный антиген, тогда как В-клетки могут распознавать протеины сахара, гликолипиды, нуклеиновые кислоты и другие виды антигенов. Эта теория была подвергнута сомнению, когда обнаружили, что CD-1 молекулы презентуют гликолипидные антигены Т-клеткам (рис. 4). Есть два вида молекул CD-1 CD-1a, CD-1b CD-1c из первой группы, CD-1d из второй группы. Обе группы презентуют антиген в человеческом организме и в организме некоторых млекопитающих, таких как кролик. У мыши антиген презентуют только молекулы СD-1 из второй группы. CD-1 молекулы похожи на молекулы МНС-I тем, что они состоят из α-цепочки, включающей три домена, которые нековалентно связаны с β2m. Анализ кристаллической структуры CD-1d показал, что α-цепочка формирует щель между первым и вторым доменами, которая уже и глубже, чем у молекул МНС 1-го класса. CD-1 антиген-связывающая щель не способна формировать водородные связи с пептидным антигеном как это делает молекула МНС 1-го класса, но хорошо подходит для гидрофобного взаимодействия. В добавок к кортикальным тимоцитам, группа I СD-1 молекул первая экспрессируется в дендритных клетках (ДК), которые являются важнейшими антигенпрезентующими (АПК) для Т-клеток. Группа II CD1 молекул наоборот, в основном находится на эпителиальных клетках, кортикальных тимоцитах и гепатоцитах, но они тоже могут быть экспрессированы на АПК таких, как ДК, макрофагах и β-клетках. При микобактериальной инфекции in vitro возрастает экспрессия CD1d молекул на поверхности мышиных ДК и макрофагов [23].
Похожие работы

... их гаплотических сочетаний у больных туберкулезом представителей русской этнической группы Челябинской области При сравнительном анализе особенностей распределения специфичностей генов HLA II класса у больных туберкулезом и здоровых представителей русской этнической группы Челябинской области были установлены следующие факты: у больных туберкулезом и здоровых пациентов были выявлены различия в ...
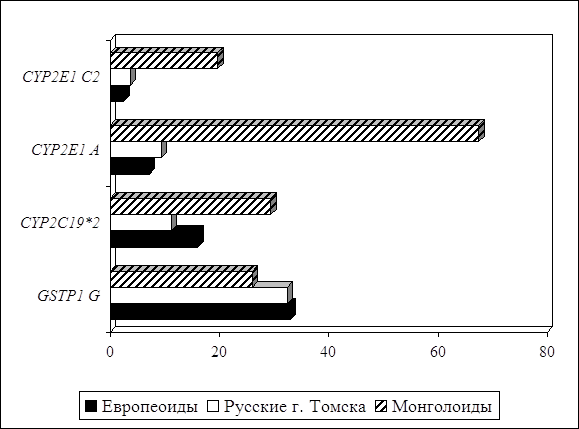
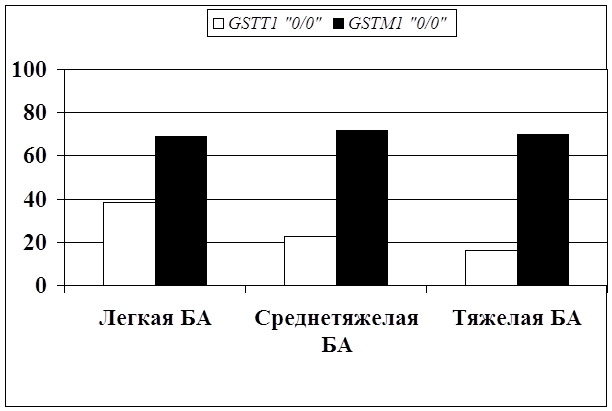
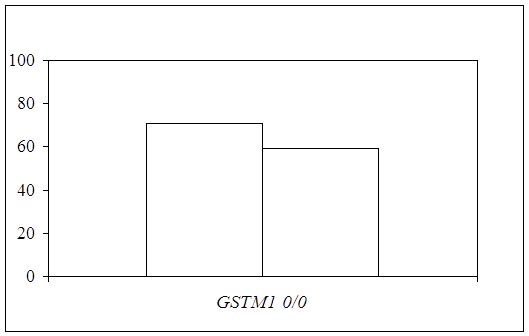
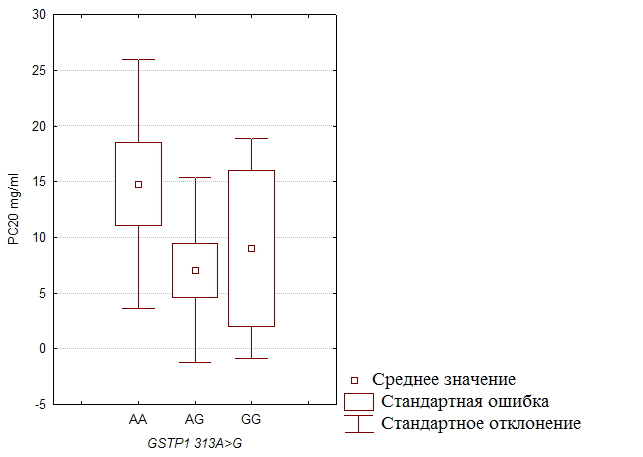
... препаратов. Установлена связь полиморфизма 313A>G гена GSTP1 с изменчивостью уровня аланинаминотрансферазы (р=0,021). 7. Выявлены различия в структуре генетической подверженности к бронхиальной астме и туберкулезу по генам ферментативной системы метаболизма ксенобиотиков: гены GSTM1, CYP2E1 и CYP2C19 связаны с бронхиальной астмой и значимыми для заболевания качественными и ...
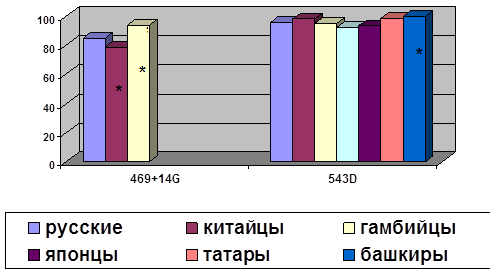
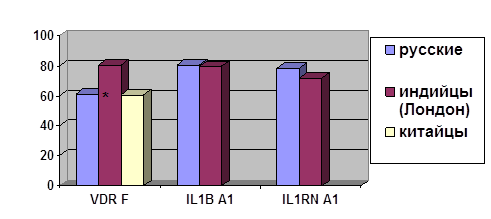
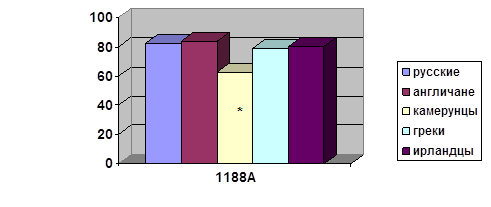
... о связи между частотой аллелей генов-кандидатов ТБ и особенностями распространения этого инфекционного заболевания. Выводы Выявлены особенности в распределении частот аллелей, изученных генов–кандидатов подвержености к туберкулезу, у русских жителей г. Томска по сравнению с другими популяциями мира. При сравнении с тувинцами показаны отличия в распределении генотипов и частот аллелей по всем ...
... фиброза и деформации лёгочной ткани. Фиброзно-кавернозный туберкулёз значительно хуже подвергается заживлению, чем кавернозный. Цирротический туберкулёз. Цирротический туберкулез лёгких характеризуется развитием в лёгочной ткани грубого, деформирующего орган склероза (цирроза), бронхоэктатических, посткавернозных типа кист полостей, эмзифематонных булл или каверн без признаков прогрессирования. ...
0 комментариев