Навигация
Ростові фактори під час децидуалізації
51123
знака
0
таблиц
0
изображений
4.5 Ростові фактори під час децидуалізації
Клітини строми у ссавців піддаються децидуальній трансформації безпосередньо перед імплантацією під впливом метаболічного сигналу з сторони бластоцисти. Перед реакцією приєднання бластоцисти до ендометрію спостерігається підвищений рівень експрессії ЕФР. Децидуальна трансформація локалізується спочатку безпосередньо біля бластоцисти, потім метаболічний сигнал епітелію передається стромі, що активує відповідну стромальну децидуальну реакцію.
Сама децидуалізація включає у себе багаточисельні зміни у стромальних клітинах. Спочатку збільшується проліферація мезенхімальних клітин, які піддаються постмітотичній диференціації і, таким чином, формують первинну децидуальну зону. Потім проліферують і диференціюються розташовані далі мезенхімальні клітини, в результаті чого утворюється вторинна децидуальна зона. Її клітини далі диференціюються і піддаються апоптозу, звільняючи таким чином місце для ембріона [10].
Серед різноманітних факторів росту, які експресуються у децидуальних клітинах у цей період – ІФР-1, рецептори до ЕФР, ТФР-α та гепарин-зв’язаного ЕФР. Наприклад, при введенні геперин-зв’яаного ЕФР у поживне середовище in vitro викликало локальну реакцію клітин ендометрію, яка полягала у підвищенні проникності судин та підсиленні процесів децидуалізації. [24]. Також є дуже важливим факт міжклітинної взаємодії між клітинами строми і ендометрію, які реалізуються сигнальними молекулами гістаміну та простагландинами. Збільшення продукції простагландину E2 модулює секрецію активатора плазміногена урокіназного типу, який експресується у стромальних клітинах ендометрію. Акумуляція активатора плазміногена і наступна судинна реакція знаходиться під контролем ЕФР. Цей ефект може інгібуватись інгібіторами транскрипції або трансляції. Ефекторні молекули родин ЕФР, інтерлейкін-1 і трансформуючі фактори росту -α та -β виявляються у гризунів на ранніх стадіях імплантації. Показано, що ген ЕФР експресується у просвіті матки миші, де знаходяться бластоцисти безпосередньо перед імплантацією. ЕФР сприяє росту бластоцисти і розростанню трофобластів in vitro. Такий же ефект спостерігається у стромальних клітинах ендометрію людини [25], [26].
Виявлена висока активність ЕФР у клітинах залозистого епітелію матки, тоді як у стромі та міометрію його рівень був набагато нижче. У свиней на 2-4-й день вагітності ЕФР виявлявся переважно в епітелії матки. На 5-й день його експресія в епітелії знижувалась, але на 6-7-й день виявлялась вже в децидуальних клітинах. Крім того, цей фактор був виявлений у трофоектодермі і у внутрішній клітинній масі доімплантаційного ембріона на 4-5-й день чи в ембріональній ендодермі та мезодермі на 5-6-й день вагітності. Передбачається, що ЕФР відіграє важливу роль у рості, міграції та адгезії клітин ендометрію, а також у формуванні міжклітинного матриксу протягом статевого циклу і на ранніх стадіях розвитку ембріона. Також рецептори ЕФР були виявлені у плаценті, що демонструє участь родини ЕФР у підтриманні вагітності. [8]
В ембріонах на ранніх стадіях розвитку продемонстровано присутність всієї родини інтерлейкінів, яка включає в себе декілька гомологічних поліпептидів та їх рецепторів. В організмі жінок поліпептиди родини ІЛ-1 були присутні у макрофагах та ендотеліальних клітинах, переважно в секреторній фазі статевого циклу [10]. На ранніх стадіях вагітност взаємодія ембріонального ІЛ-1 та рецепторів ІЛ-1 в ендометрії може забезпечувати впізнання та імплантацію ембріона. [1], [2].
ІЛ-1 та ТФР-β відіграють суттєву роль в імплантаційних процесах за рахунок регуляції експресії стромою ендометрію тканинних інгібіторів металлопротеїназ –1 та –3, а також колагенази IV типу, які є дуже важливі при інвазії трофобласта. Зниження синтезу колагена IV типу в стромі матки починається в середині секреторної фази і продовжується після імплантації. Рахується, що зменшення в’язкості цього типу колагену – головний фактор, який сприяє розвитку набряку у зовнішньоклітинному просторі при імплантації бластоцисти. Прогестерон інгібував стимулюючий ефект ІЛ-1 у відношенні металопротеїнази-3 у секреторній фазі in vivo та in vitro. Відомо, що подавлення активності металопротеїназ є необхідним для зберігання стабільності тканин при інвазійних процесах при імплантації та розвитку плаценти. ЕФР та ФРФ, навпаки, підвищували рівень протеолітичних ферментів, які продукуються ендометріальними стромальними клітинами у процесі децидуалізації in vitro , але не впливали на активність їх інгібіторів. [9]
4.6 Ростові фактори під час імплантації
Основну роль у проліферації клітинних компонентів залозистого епітелію ендометрію, строми, гладеньких м’язів та ендотелію судин відіграють специфічно експресовані під час проліферативної фази циклу ЕФР, ІФР-1 та ІФР-2, ФРФ.
Вважається, що інсуліноподібні фактори росту стимулюють мітоз і диференціацію клітин ендометрію протягом статевого циклу і на ранній стадії вагітності, є мітогенними для клітин строми та залозистого епітелію. ІФР-1 індукує багаточисельні локальні відповіді у клітинах ендометрію – такі, як підвищення локальної проникності судин, децидуалізацію та експресію деяких генів. Під час імплантації на стадії інвазії трофобласта ІФР-1 та ІФР-2 виявляють різні по локалізації та часовим характеристикам профілі експресії. Так, ІФР-1 експресувався у стромі та залозистому епітелії – тобто на ділянках, які є початковими на етапах прикріплення та інвазії трофобласта. Велика кількість ІФР-2 була виявлена в частині апікальних клітин, тоді як у базальному шарі на латеральних плазматичних мембранах більше виявляється ІФР-1. Це свідчить про вирішальну роль ІФР-2 у процесах саме адгезії. Тварини, у яких порушений ген ІФР-1, проявляли знижену фертильність. В різних групах клітин трофобласту виявляється знижений рівень ІФР-1 і навпаки, підвищений рівень ІФР-2. [10]
При вивченні родини ІФР-зв’язуючих білків встановлено, експресія ІФРСБ-1 знаходиться під контролем прогестерону, а сам ІФРСБ-1 виступає в ролі білка, який захищає материнський організм від надлишкової інвазії трофобласта і вже на 7-10-й день циклу ці білкі виявляються в децидуальних клітинах на ділянці імплантації.. [19]
Неопосередкована дія 17-β-естрадіолу на проліферацію епітеліальних клітин ендометрію проявляється у впливі естрадіолу на секрецію ІФР-1 стромальними клітинами ендометрію. Одночасно виявлені стимулюючі ефекти інсуліну на проліферацію клітин ендометрію, які здійснюються через активацію стромальних клітин. [11], [27].
5. Список літератури
1. Гьюдай Линда С. Имплантирующаяся оплодотворённая яйцеклетка и материнский организм // Проблемы эндокринологии, 1999; Т.5. С.30-32
2. Чернуха Г.Е., Сметник В.П., Роль факторов роста в функции репродуктивной системы // Проблемы эндокринологии, 1996, Т.2. С.8-13
3. Фильченков А.А., Стойка Р.С., Быкорез А.И. Трансформирующие факторы роста, 1994, 291 С.
4. Monniaux D., Monget P., Besnard N. Growth factors and antral follicular development in domestic ruminants. // Theriogenology.-1997. Vol.47. №1. P. 3-12
5. Abigail L. Fowden. Endocrine regulation of fetal growth // Reproduction. Fertility. Development // 1995. Vol.7. P.351-363
6. Aplin J.D. Adhesion molecules in implantation // Reviewes of reproduction. 1997. Vol.2. P.84-93
7. Bass K., Morrish D., Roth I. Human cytotrophoblast invasion is up-regulated by EGF:evidence that paracrine factors modify this process // Developmental biology.- 1994.- Vol.164. №2.- P.550-556
8. Buyalos R.P., Cai X. Preimplantation embryo development enhanced by Epidermal Growth Factor // Journal of Assisted Reproduction and Genetic.-1994. Vol.11. №1. P.33-37
9. Carson D.D., Bangchi I., Dey S.K. Embryo implantation // Developmental biology. 2000. Vol.223. P.217-237
10. Carver J., Martin K., Spyropoulou I. An in vitro model for stromal invasion during implantation of the human blastocyst // Human reproduction. 2005. Vol.18. P.283-290
11. Davis J.S., May J.V., Keel B.A. Mechanisms of hormone and growth factors action in the bovine corpus luteum //Theriogenology.- 1996.- Vol. 45,№ 7.- Р.1351-1380
12. Derrar N., Price C.A., Sirard M.A., Effect of growth factors and co-culture with ovarian medulla on the activation of primordial follicles in explants of bovine ovarian cortex // Theriogenology. 2000. Vol.54. №4. P.587-598
13. Duque P., Gómez E., Díaz E. et all, Use of two replacements of serum during bovine embryo culture in vitro // Theriogenology. 2005. Vol.59. №3-4. P.889-899.
14. Fujimoto J., Sakaghuchi H., Hirose R., Tamaya T. Significance of sex steroids in roles of cadherin subfamily and its related proteins in the uterine endometrium and placenta.// Horm.Res. 1998. Vol.50. P.30-36
15. Glister C., Groome N., Khight. Oocyte-mediated suppression of FSH and IGF-induced secretion of steroids and inhibin-related proteins by bovine granulose cells in vitro: possible role of TGF-α .// Biology of reproduction. 2003. Vol.68. P.758-765
16. Guler A., Poulin N., Mermillod P., Effect of growth factor EGF and IGF-I and estradiol on in vitro maturation of sheep oocytes // Theriogenology.2000. Vol. 54. №2. P.209-218
17. Iwakura Y. Mechnism of blastocyst formation of the mouse embryo // Development // Growth and Differentiation. 1989. Vol.31. №6. P.523-529
18. Kokawa K., Shikone T., Nakano R., Apoptosis in the human uterine endometrium during the menstrual cycle.// J.Clin.Endocrinol.Metab, 1996. Vol.81. P.4144-4147
19. Lackey B.R., Gray S.L., Henricks D.M. Physiological basis of use of insulin-like growth factors in reproductive applications: A Review // Theriogenology.- 2000.-Vol.53, № 5.- P.1147-1156
20. Lee K.Y., De Mayo F.J Animal models of implantation // Reproduction. 2004.Vol.128. P.679-695
21. Mann G.E., Green M.P., Sinclair K.D., Effects of circulating progesterone and insulin on early embryo development in beef heifers //Animal Reproduction Science. 2003. Vol.79. №1-2/ P.71-79
22. Merriman J.A., Whittingham D.G. and Carroll J., The effect of follicle stimulating hormone and epidermal growth factor on the developmental capacity of in-vitro matnrated mouse oocytes // Human Reproduction. 1998. Vol.13. № 3. P.690-695
23. Sharkey A. Cytokines and implantation // Reviewes of reproduction. 1998. Vol.2. P.84-93
24. Sirisathein S., Hernandes-Fonseca H.J., Influences of Epidermal Growth Factor and Insulin Growth factor-I on bovine blastocyst development in vitro // Animal Reproduction Science. 2003. Vol.77. №1-2/ P.21-32
25. Spencer T.E., Burghardt., Johnson G.A., Bazer F.W. Conceptus signals for establishment and maintenance of pregnancy // Animal Reproduction Science. 2004. Vol.82-83. P.537-550
26. Taga M., Suginami H., Cell adhesion and reproduction. // Horm.Res. 1998. Vol.50. P.2-6
27. Ueda O., Yorozu K., Kamada N. Possible expansion of "window of implantation" in pseudopregnant mice: time of implantation of embryos at different stages of developmental transferred into the same recipient // Biology of reproduction. 2003. Vol.69. P.1085-1090
28. Wang Y., Rippstein P. U., Tsang B. K. . Role and Gonadotrophic Regulation of X-Linked Inhibitor of Apoptosis Protein Expression During Rat Ovarian Follicular Development In Vitro//Biology of Reproduction. 2003. Vol. 68. №2. P. 610 - 619.
Похожие работы
... кількості мітохондрій в період оогенезу. У багатьох тварин розмноження мітохондрій відбувається в основному до початку вітелогенезу. Наприклад, у жаби в період вітелогенезу і на ранніх стадіях ембріогенезу мітохондрії практично не діляться, тобто інтенсивне розмноження іде в превітелогенних ооцитах [Айзенштадт, 1984]. Таким чином, активна реплікація мітохондріальної ДНК в превітелогенних ооцитах ...


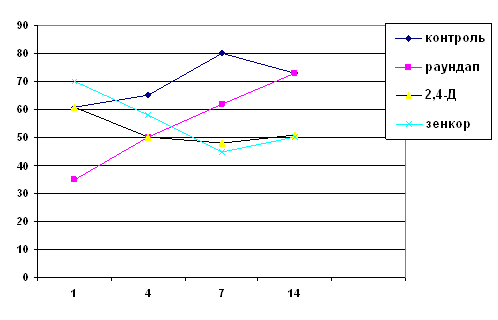
... впливу фосфорорганічного пестициду діазинону дозоволило авторам роботи зробити висновок про значні морфологічні та фізіологічні зміни у Melanotaenia duboulayi, що виражались у гіпертрофії та гіперплазії зябер, закупорці кровоносних судин, потовщенні епітелію зябрових пелюсток, посиленні секреції слизу. При хронічному отруєнні похідними дихлорфеноксиоцтової кислоти у риб спостерігається ...
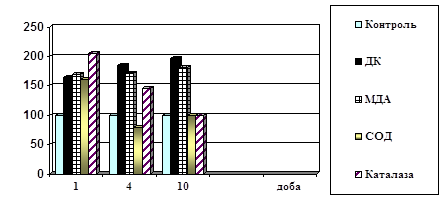
... ів України. – Київ, 2003. – С.312. (Внесок дисертанта: проведено визначення каталази в крові хворих і підготовлено матеріал до друку) АНОТАЦІЯ Поліянц І.В. Патофізіологічні механізми пневмонії на різних етапах її розвитку. – Рукопис. Дисертація на здобуття наукового ступеня кандидата медичних наук за фахом 14.03.04 – патологічна фізіологія. Одеський державний медичний університет МОЗ Укра ...
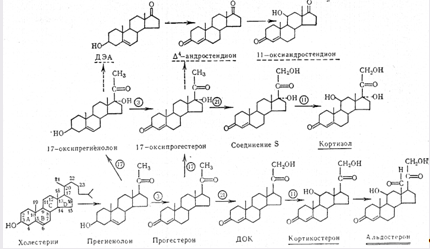

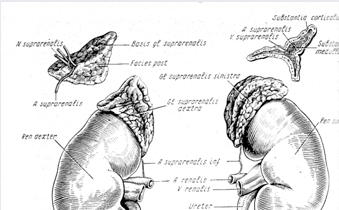
... контролюють гормоноутворення в кожній із зон, різноманітні. Але ж при всій своїй відносній самостійності окремих зон, вони зберігають відому структурну єдність та функціональну взаємодію, тому кора наднирників є цілісною системою. Дві частини наднирників продукують гормони, що різко відрізняються за хімічним складом і фізіологічною дією в організмі, регуляція їх відбувається окремо. Кіркова і ...
0 комментариев