Навигация
3.3 Процеси адгезії
Маркерами ендометріальної функціональності та готовності ендометрію до імплантації є молекули адгезії. Адгезія - це процес прикріплення бластоцисти до поверхні матки. Цей процес опосередкований активністю цитоскелету, специфічних білків адгезії та їх рецепторів, що забезпечує адекватну локалізацію бластоцисти і активацію внутрішньоклітинних сигнальних біохімічних шляхів, що супроводжує клітинну активацію. Всі молекули, які приймають участь в процесах адгезії, є клітинними поверхневими рецепторними білками. Є 4 основних групи таких молекул: інтегріни, кадгеріни, селектини та супергруппа іммуноглобілінов [27], [14].
Інтегріни взаємодіють з різними лігандами, включаючи глікопротеїни зовнішньоклітинного матриксу та молекули на клітинних мембранах. Вони полегшують міграцію та прикріплення клітин до матриксу, опосередковують міжклітинні взаємодії та передачу сигналів. Ендометрій відноситься до тканин з високим рівнем експресії інтегрінів, які відіграють значну роль у процесах запліднення, імплантації та розвитку плаценти. Рахується, що присутність трьох типів цих сполук – a1b1, a4b1, avb3 - тільки протягом 20-24 дня циклу полегшує імунологічне розпізнавання ембріона та забезпечує позитивний результат імплантації. Цікаво, що додавання деяких молекул адгезії у культуральне середовище підвищувало якість ембріонів та збільшувало частоту їх наступної імплантації при трансплантації [6], [27].
Клітинна адгезія - важливий регулятор апоптозу. Індукція апоптозу за рахунок порушення взаємодії клітин із субстратом особливо добре проявляється в ендотеліальних та епітеліальних клітинах. Показано, що зміни у структірі інтегринів можуть вплинути на розвиток апоптоза.
3.4 Апоптоз на ранніх стадіях ембріонального розвитку
Це запрограмована клітинна смерть, яка контролюється генетично та характеризується активацією протеолітичних ферментів (каспаз), специфічною фрагментацією ДНК, розривами плазматичної мембрани та розпадом клітини на апоптичні тільця. Саме апоптозом контролюються характерні морофологічні зміни, які визначають готовність ендометрію до імплантації бластоцисти. Встановлено, що апоптоз розвивається у специфічних популяціях клітин ендометрію протягом усього статевого циклу. В ранньому проліферуючому ендометрію апоптозу підлягають клітини функціонального шару, у пізню проліферативну фазу і до самої середини секреторної фази ознак апоптозу в ендометрії не спостерігається. [18]
На початку пізньої секреторної фази апоптоз знову спостерігається у стромальних клітинах ендометрію. Початку апоптозу в залозах і стромі передує зниження концентрації 17-β естрадіолу та прогестерону у сироватці крові. Зниження кількості рецепторів до естрогену і прогестерону в матковому епітелії обернено корелює з ростом апоптичного індекса в цих тканинах. Клітини базального шару не піддаються апоптозу ні в одну з стадій естрального циклу. Продемонстровано, що можна експериментально моделювати запуск процесів апоптозу в ендометрії тварин in vitro шляхом вилучення стероїдних гормонів. Встановлено, що активація апоптоза в ендометрії корелює із змінами в концентрації 17-β естрадіолу та прогестерону у сироватці крові під час циклу, концентрацією клітинних рецепторів до естрогенам і прогестерону, а також з кількісними характеристиками проліферативних і секреторних змін в епітеліальних і стромальних клітинах ендометрію.
Показано участь апоптичних змін ендометрію в ході нормального процесу імплантації мишачих ембріонів. [28].
3.5 Роль трофобласта у процесах інвазії
Після прикріплення бластоциста починає зморщуватись і вкорочуватись та в інвазивну стадію вступає більш ущільненою. На цій стадії головну роль відіграє трофобласт, оскільки саме цією частиною бластоциста занурюється в строму. Занурення відбувається з обох боків – всередині знаходиться частина ембріобласту. Тобто трофобластична частина керує всім процесом інвазії взагалі. Бластоциста в такому стані набуває біполярну вісь симетрії – таку ж саму орієнтацію приймають і клітини строми в тому місці, куди імплантується бластоциста – очевидно, що регуляція є двосторонньою і взаємною.
При цитологічних дослідження (фарбуванні) тотальних препаратів добре видно короткі хрестоподібні структури волокон актину трофобласту поверх більш темних волокон стромального актину, розташованих нижче. Існують дані, які підтверджують наявність волокон актину та цитокератину в самих клітинах трофобласту на останніх стадіях перед інвазією, що може служити своєрідним “якорем” для укорінення в строму. Трофобласт проростає в строму так глибоко, що покривний епітелій повністю змикається над ним. [7]
4. Роль білкових ростових факторів у встановленні вагітності
4.1 Ростові фактори – регулятори ембріонального розвитку
При підготовці материнського організму до імплантації в якості локальних клітинних медіаторів дії стероїдів, які задіяні в циклічних змінах ендометрію, виступають ростові фактори. В доімплантаційний період вони присутні в ендометріальній тканині в дуже значних кількостях. Епідермальний фактор росту (ЕФР), інсуліноподібні фактори росту (ІФР-1 і 2), фактор росту фібробластів (ФРФ), фактори росту родини трансформуючих факторів росту (ТФР) активують мітотичну активність і властивість до диференціації клітин ендометрію, що підвищує сприйнятливість його до бластоцисти, яка імплантується під час “вікна імплантації”. Дією факторів росту опосередковуються специфічні зміни кількісного і якісного складу субпопуляцій лейкоцитів, які обумовлюють адекватну материнську імуносупресію та ендометріальну відповідь на занурення трофобласта.
Джерелом синтезу та секреції більшості ростових факторів і цитокінів є епітеліальні клітини, макрофаги та лімфоцити. Експериментально доведено, що синтез цитокінів нативними клітинами-кіллерами знаходиться під контролем прогестерону. Фактори росту і цитокіни являють собою декілька родин пептидів, які залучені у паракринні, інтракринні та аутокринні механізми регуляції клітинних реакцій за рахунок зв’язування із специфічними рецепторами клітинної поверхні [3], [23].
ссавець вагітність імплантація
Похожие работы
... кількості мітохондрій в період оогенезу. У багатьох тварин розмноження мітохондрій відбувається в основному до початку вітелогенезу. Наприклад, у жаби в період вітелогенезу і на ранніх стадіях ембріогенезу мітохондрії практично не діляться, тобто інтенсивне розмноження іде в превітелогенних ооцитах [Айзенштадт, 1984]. Таким чином, активна реплікація мітохондріальної ДНК в превітелогенних ооцитах ...


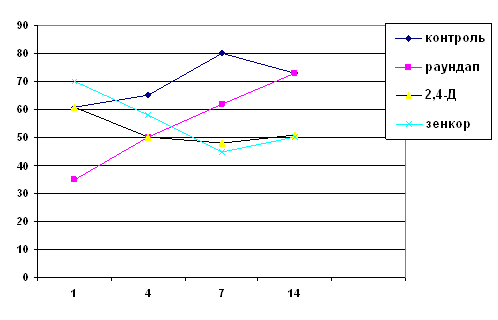
... впливу фосфорорганічного пестициду діазинону дозоволило авторам роботи зробити висновок про значні морфологічні та фізіологічні зміни у Melanotaenia duboulayi, що виражались у гіпертрофії та гіперплазії зябер, закупорці кровоносних судин, потовщенні епітелію зябрових пелюсток, посиленні секреції слизу. При хронічному отруєнні похідними дихлорфеноксиоцтової кислоти у риб спостерігається ...
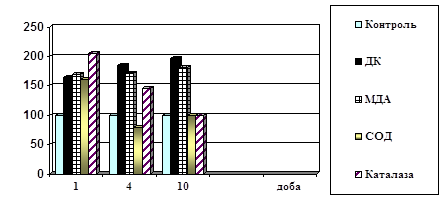
... ів України. – Київ, 2003. – С.312. (Внесок дисертанта: проведено визначення каталази в крові хворих і підготовлено матеріал до друку) АНОТАЦІЯ Поліянц І.В. Патофізіологічні механізми пневмонії на різних етапах її розвитку. – Рукопис. Дисертація на здобуття наукового ступеня кандидата медичних наук за фахом 14.03.04 – патологічна фізіологія. Одеський державний медичний університет МОЗ Укра ...
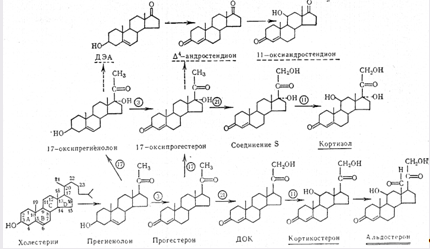

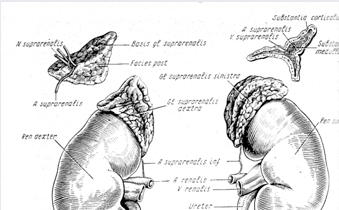
... контролюють гормоноутворення в кожній із зон, різноманітні. Але ж при всій своїй відносній самостійності окремих зон, вони зберігають відому структурну єдність та функціональну взаємодію, тому кора наднирників є цілісною системою. Дві частини наднирників продукують гормони, що різко відрізняються за хімічним складом і фізіологічною дією в організмі, регуляція їх відбувається окремо. Кіркова і ...
0 комментариев