Навигация
Передвижение воды по растению
274314
знаков
4
таблицы
0
изображений
4. Передвижение воды по растению.
При передвижении по клеткам паренхимы корня вода обогащается минеральными веществами и в таком составе попадает в клетки ксилемы, скелетной основой которой являются сосуды и трахеиды. Сосуды, у которых нет протоплазмы, обладают высокой сосущей силой, пропорциональной осмотическому давлению содержащегося в них раствора.
Находящаяся в сосудах и трахеидах вода имеет форму тончайших нитей, которые своими верхними концами как бы подвешены к испаряющим клеткам листьев, а нижними концами упираются в паренхимные клетки корня. Для того, чтобы вода передвигалась вверх, необходимо, чтобы испаряющие клетки обладали достаточной величиной сосущей силы. В отсутствие этого условия возникает ток воды в сосудах в обратном направлении.
За счет того, что в атмосфере почти всегда содержится воды меньше, чем в растении, определяется явление отрицательного водного потенциала и, следовательно, сосущей силы атмосферы. Сосущая сила в испаряющих клетках достигает 2-4 тысяч килопаскаль.
Удерживание воды в сосудах ксилемы в виде нитей обуславливается силами когезии и адгезии.
Когезия - это прочное сцепление молекул воды между собой.
Адгезия - это прилипание молекул воды к гидрофильным стенкам клеток ксилемы.
При передвижении воды в клетках ксилемы возникает электрический заряд вдоль поверхностей раздела клеток (по мембранам). Электроосмос определяется наличием проницаемых мембран с системой пор разной величины и постоянной диффузии электролитов, которыми и является передвигаемая по ксилеме пасока (вода и растворенные в ней минеральные вещества).
Состав пасоки сильно варьирует в зависимости от вида растения и фазы его вегетации и фазы органогенеза. Пасока однолетнего травянистого растения и многолетнего древесного растения безусловно сильно отличаются друг от друга, так же как и пасока у одного и того же растения весной, летом и осенью. У ряда древесных растений человек использует весеннюю пасоку в своем питании (березовый сок, кленовый сок). Пасока, выделяющаяся при гуттации, имеет в своем составе очень мало минеральных веществ и сахаров, поскольку происходит их естественная фильтрация при прохождении пасоки через эпитему (ткань, выстилающую воздушную полость гидатоды).
Транспирация.
Завершающей частью водного обмена растений является транспирация, или испарение воды листьями, то есть верхний двигатель тока воды в растении. Это явление с физической стороны представляет собой процесс перехода воды в парообразное состояние и диффузию образовавшегося пара в окружающее пространство.
Транспирация выполняет в растении следующие основные функции:
это верхний двигатель тока воды,
это защита от перегрева,
это нормализация функционирования коллоидных систем клеток листа.
Транспирация характеризуется следующими показателями: интенсивностью, продуктивностью и коэффициентом.
Интенсивность транспирации - это количество воды, испаряемой растением с единицы листовой поверхности в единицу времени. Выражается формулой:
Тр= DС г Н2О _
еr м2.1час,
где Тр - интенсивность транспирации, DС - градиент концентрации водяного пара между транспирирующей поверхностью и окружающим воздухом, еr - сумма диффузионных сопротивлений листа (устьичного, кутикулярного и сопротивления пограничного слоя).
Сопротивление пограничного слоя зависит от ветра, при отсутствии ветра оно максимально, чем больше ветер, тем оно меньше.
Устьичное диффузионное сопротивление зависит от степени открытия устьиц.
Кутикулярное диффузионное сопротивление зависит от толщины кутикулярного слоя, чем она больше, тем больше сопротивление.
Продуктивность транспирации - это количество созданного сухого вещества на 1 кг транспирированной воды. В среднем эта величина равна 3 г/1 кг воды.
Транспирационный коэффициент показывает сколько воды растение затрачивает на построение единицы сухого вещества, т.е. этот показатель является величиной, обратной продуктивности транспирации и в среднем равен 300, т.е. на производство 1 тонны урожая затрачивается 300 тонн воды.
Очень важным моментом в процессе транспирации является действие абиотических факторов окружающей среды: влажности атмосферного воздуха и температуры воздуха.
Чем менее влажен атмосферный воздух, т.е. чем меньше его водный потенциал, тем интенсивнее будет идти транспирация. При 100% влажности воздуха его водный потенциал равен нулю. Уже при снижении влажности воздуха на 1-2% его водный потенциал становится отрицательной величиной, а при снижении влажности воздуха до 50% показатель водного потенциала выражается отрицательной величиной порядка 2-3 сотен бар в зависимости от температуры воздуха. При этом в клетках листьев показатель водного потенциала, как правило, выше нуля, поэтому диффундирование воды из межклетников в атмосферу наблюдается почти всегда.
Чем выше температура воздуха, тем выше будет и температура листа, при этом температура внутри клеток листа может быть на 10оС выше, чем в атмосфере. Происходит нагрев воды, находящейся в листе, что также способствует процессу испарения.
Регулировка транспирация происходит в растении по двум механизмам:
устьичная регуляция,
внеустьичная регуляция.
Наиболее существенной является устьичная регуляция, которая определяется как некоторыми физическими закономерностями, так и влиянием ряда факторов внешней среды и внутренней биохимией клеток листа.
С физической точки зрения основой испарения из устьица является физический механизм испарения с ограниченных поверхностей очень маленькой площади. При этом имеет значение величина снижения упругости водяного пара (F-f) и расстояние (l), на протяжении которого поддерживается эта разница, которая определяет градиент дефицита насыщения.
F-f
D D = - --------
l
При этом скорость испарения V будет пропорциональна градиенту насыщения, а А - постоянная, определяемая прочими условиями, влияющими на скорость испарения:
F-f
V = А - --------
l
Поскольку речь идет об ограниченных поверхностях (устьице), то краевое испарение за счет меньшей величины l2 будет выше, чем в центре, т.е.:
F-f F-f
--- - > - -----
l2 l1
Применительно к испарению с площади круга формула скорости испарения принимает вид
V = k R2,
где k - значение всех прочих факторов, определяющих скорость испарения, а R - радиус круга.
При испарении с малых поверхностей, когда доля участия краевого испарения значительна, формула видоизменяется в
V = k Rn,
где n - положительное число между 1 и 2, т.е.2 >n>1. В случае малых площадей, таких как отверстие устьичной щели, n становится равным 1. Таким образом определяющим становится фактор k, т.е. суммарное значение факторов окружающей среды и суммарное количество устьиц на листе.
В устьичной транспирации ведущими факторами являются:
количество устьиц на единицу листовой поверхности,
форма листа (чем более причудлива форма листа, тем больше его площадь, а, значит, и количество устьиц),
наличие ионов К+ (чем выше концентрация, тем больший приток воды в замыкающие клетки устьица и тем шире устьичная щель),
наличие абсцизовой кислоты (чем выше концентрация этого гормона старения, тем меньше раскрытие устьица) (пример - мутант томата wilty),
концентрация углекислого газа в подустьичной полости (чем ниже концентрация, т.е. меньше 0,03%, находящихся в воздухе, тем больший приток воды в замыкающие клетки устьица и тем шире устьичная щель),
наличие солнечного света (на свету крахмал превращается в простые сахара, т.е. концентрация клеточного сока выше, поэтому наблюдается больший приток воды в замыкающие клетки устьица и раскрытие устьичной щели),
наличие и скорость ветра (непосредственно к испаряющей поверхности прилегает слой воздуха, в котором водяной пар постепенно испаряется далее в атмосферу, при этом в безветренную погоду скорость испарения выражается линейной зависимостью между дефицитом насыщения воздуха и расстоянием от испаряющей поверхности. Однако, при наличии ветра, который "сдувает" испаряющиеся молекулы воды, происходит увеличение дефицита насыщения воздуха. Возле поверхности листа сохраняется лишь небольшой ламинарный слой (dS), сохраняющийся и при сильном ветре, где можно наблюдать линейную зависимость дефицита насыщения от расстояния).
Внеустьичная транспирация определяется количеством и размерами межклеточных пор в кутикуле листа. Радиус клеточных пор очень мал, составляет около 100-200 Ао, т.е. около 0,00001мм, однако в листе имеющем много кутикулярных пор скорость испарения снижается достаточно значительно, иногда почти в два раза.
Различают три вида движения устьиц (закрытие и открытие устьиц):
фотоактивные (под действием солнечного света),
гидроактивные (при потере воды),
гидропассивные (при дожде из-за набухания клеток эпидермиса и сдавливания устьичных клеток).
Суточный ход транспирации у всех растений определяется максимальной транспирацией в утренние часы и минимальной - в полуденные. При этом весьма существенное значение имеют и такие факторы, как температура почвы и воздуха, влажность почвы и воздуха, интенсивность солнечного излучения, наличие ветра.
Сезонный ход транспирации у многолетних растений определяется фазами развития растения.
Водный баланс в растении.
Водный баланс в растении поддерживается тогда, когда скорость поглощения воды равна скорости ее испарения. Обычно водный баланс в растении меняется в течение суток, при этом он зависит от уровня агротехники при выращивании растений, т.е. от уровня орошения и удобрения. Несбалансированность поступления и испарения воды проявляется в наличии водного дефицита, который наблюдается, как правило, у растений днем и отсутствует ночью.
В практике сельского хозяйства используются приемы, снижающие водный дефицит у растений: Использование освежительных поливов, Использование антитранспирантов.
Антитранспиранты делятся на две разновидности:
вещества, вызывающие закрытие устьиц (абсцизовая кислота, фенилмеркурацетат),
вещества, образующие пленки на листьях (полиэтилен, латекс).
Лекция 10-12
Тема: Фотосинтез.
Дополнительная литература:
Н.Н. Овчинников, Н.М. Шиханова. Фотосинтез. М., 1972
Пигменты пластид зеленых растений и методика их исследований. Под ред. Сапожникова. Изд-во "Наука", М. - Л., 1964.
И.А. Шульгин. Солнечная радиация и растение. Изд-во "Гидромет", Л., 1967.
Ю.С. Насыров. Фотосинтез и генетика хлоропластов. Изд-во "Наука", М., 1975.
Вопросы к теме:
Общая характеристика фотосинтеза.
Лист как орган фотосинтеза.
Пластиды (хлоропласты, хромопласты), хлорофиллы, каротиноиды.
Световая фаза фотосинтеза.
Фотосистема 1.
Фотосистема П.
Фотолиз воды или реакция Хилла.
Темновая фаза фотосинтеза или цикл Кальвина (С-3 путь фотосинтеза).
С-4 путь фотосинтеза.
САМ-фотосинтез.
Фотодыхание.
Характеристика основных показателей фотосинтеза: интенсивности и продуктивности.
Усвоение растением фотосинтетически активной радиации.
Условия, влияющие на интенсивность и продуктивность фотосинтеза.
Пути повышения продуктивности фотосинтеза.
Суточный ход фотосинтеза у светолюбивых и теневыносливых растений.
Роль зеленых растений в природе.
Общая характеристика фотосинтеза.
Жизнь на нашей планете обеспечивается энергией фотонов, содержащейся в солнечном излучении. Эта энергия (кванты солнечного света - физическая форма энергии) поглощается фотоавтотрофными организмами - в подавляющем большинстве являющимися растениями. Поглощение или абсорбция энергии осуществляется специфическими молекулами - пигментами, которые способны ее превращать в энергию химических связей. В дальнейшем поглощенная и преобразованная энергия тратится автотрофами на присоединение молекулы углекислого газа к органической молекуле, то есть на синтез органического вещества из неорганического. Общеизвестна реакция фотосинтеза:
6СО2 + 6 Н2О = С6 Н 12О6 + 6О2
Фотосинтез состоит из двух сопряженных процессов:
Окисления воды до кислорода
Восстановления углекислого газа водородом воды до полисахаридов.
Многие годы предполагали, что кислород освобождается из молекулы углекислого газа, но изучение процесса фотосинтеза у микроорганизмов и выявление у них способности использовать в качестве доноров электронов не воду, а другие водородсодержащие вещества, позволило установить, что кислород получается в результате разложения воды.
Оказалось, что процесс фотосинтеза состоит из двух фаз:
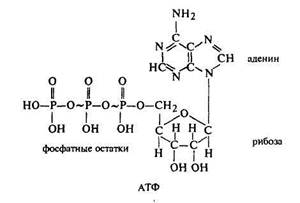
световой, в которой разлагается вода под действием энергии солнца, абсорбированной пигментами, и происходит запасание энергии в виде химических связей в макроэргических молекулах (АТФ и НАДФ),
темновой, в которой происходит собственно синтез органических веществ (глюкозы, а затем крахмала) из углекислого газа за счет использования энергии, накопившейся в световой фазе.
Лист как орган фотосинтеза.
Лист растения - это основной орган растения, где проходит процесс фотосинтеза. Поскольку в основном лист покрыт малопроницаемой для газов кутикулой, то поступление СО2 в ткани идет через устьица, а в тканях - через сильно разветвленную сеть межклеточных воздухоносных каналов.
К верхней стороне листа прилегает палисадная паренхима, клетки которой расположены перпендикулярно, плотно соприкасаются друг с другом и содержат много хлоропластов. Эта палисадная паренхима и является основной ассимиляционной тканью. К нижнему эпидермису прилегает губчатая паренхима с рыхло расположенными клетками и межклетниками. Кроме того, весь лист пронизан жилками, по которым идет перенос веды, минеральных ионов и ассимилятов.
В палисадной паренхиме нет ни одной клетки, которая отстояла бы от ближайшей к ней жилки дальше, чем на несколько клеточных диаметров.
Пройдя сквозь устьичный барьер, атмосферный СО2 растворяется в воде, гидратируется и превращается в угольную кислоту, а затем диссоциирует до бикарбонат-ионов (НСО3+), запас которых и служит резервом потенциального СО2 для использования в фотосинтезе.
Поскольку основной тканью, поглощающей энергию солнца, является палисадная паренхима, содержащая максимальное количество хлоропластов, то, зная соотношение между площадью листа и площадью поверхности хлоропластов, можно приблизительно определить и поглощающую способность посевов. Так, на 1 га посева в среднем приходится 5 га листовой поверхности, то есть 1000 га поверхности хлоропластов, так как 1 см2 листовой поверхности соответствует 200 см2 поверхности хлоропластов. При этом площадь поверхности межклетников, испаряющих воду составляет 50 га. В этом проявляется общебиологический закон - создание внутренних рабочих поверхностей при сравнительно малых наружных испаряющих площадях за счет затрат небольших количеств материала.
В зависимости от условий обитания растений (засушливый или избыточно влажный климат, тропический климат с чрезмерной интенсивностью солнечного излучения) в строении листьев могут наблюдаться те или иные морфологические или биохимические особенности, однако общие принципы строения листа сохраняются.
Пластиды (хлоропласты, хромопласты), хлорофиллы, каротиноиды.
В растениях встречается три типа пластид, которые делятся в зависимости от типа пигментов, входящих в их состав:
хлоропласты,
хромопласты,
лейкопласты.
Для процесса фотосинтеза важнейшую роль играют хлоропласты, содержащие хлорофиллы. Хромопласты или отдельные группы каротиноидов могут участвовать в процессе фотосинтеза, однако их роль более вспомогательная. Однако, встречаются растения с преобладанием хромопластов (японская слива, декоративные краснолистные формы), которые самостоятельно осуществляют процесс фотосинтеза.
Строение хлоропласта - двойная мембрана, отделяющая хлоропласт от цитоплазмы, фотосинтетические мембраны - тилакоиды стромы и тилакоиды гран, наличие участков ДНК, способность к цитоплазматическому наследованию. Внутренние части полости тилакоидов гран и межгранальные тилакоиды - это единая замкнутая фотосинтетическая внутримембранная полость, объединенная в единую фотоэнергетическую систему хлоропласта.
Грана хлоропласта состоит из 10-30 тилакоидов, а всего в хлоропласте 100-150 гран, таким образом поверхность фотосинтетических мембран тилакоидов в 10 раз превышает поверхность самого хлоропласта.
Особая роль отводится концевым тилакоидам граны, которые, будучи селективным фильтром, предохраняют грану от излишнего облучения или подают сигнал на изменение ориентации оси граны. При оптимальных условиях освещения оси гран обычно направлены радиально к более выпуклой стороне хлоропласта.
Функция хлоропласта - осуществление процесса световой фазы фотосинтеза и накопление энергии в виде макроэргических молекул (АТФ и НАДФ восстановленного).
Свойства хлоропластов - способность к перемещению внутри клетки под воздействием условий освещенности и концентрации углекислого газа. Передвижение хлоропластов по клетке называется фототаксисом или хемотаксисом хлоропластов в зависимости от причины, вызывающей это передвижение. При умеренном освещении хлоропласты выстраиваются таким образом, чтобы на них попадало максимальное количество света, а при избыточном освещении выстраиваются вдоль падающих солнечных лучей. Такое расположение хлоропластов называется парастрофией. Ночью хлоропласты выстраиваются в положении апострофии.
Хромопласты придают желтую, оранжевую, красную окраску лепесткам, плодам, листьям, так как содержат большое количество специфических каротиноидов, обладающих тем или иным оттенком окраски. Хромопласты функционально дополняют деятельность хлоропластов, кроме того выполняют функцию привлечения насекомых-опылителей, животных-распространителей семян.
В состав фотосинтетических мембран (тилакоидов) входят специфические фотосинтетические пигменты - хлорофиллы и каротиноиды - погруженные в эти мембраны.
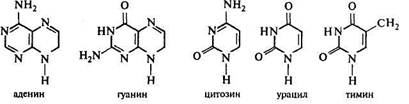
Хлорофиллы делятся на четыре разновидности: а, b, c, d. Это органические соединения, содержащие 4 пиррольных кольца, связанных атомами магния и имеющими зеленую окраску. Отличаются между собой хлорофиллы по молекулярной массе:
а - имеет молекулярную массу 893 и включает фитоловый и метиловый остаток,
b - имеет молекулярную массу 907 и включает фитоловый и метиловый остаток,
с - включает только метиловый остаток,
d - имеет молекулярную массу 891 и близок к протохлорофиллу.
У высших растений встречаются в основном хлорофиллы а и b, а у водорослей - а и с или а и d.
Хлорофилл впервые был выделен в 1818 году, к 1940 году была расшифрована его структура, а в 1960 году осуществили синтез хлорофилла. Хлорофиллы - это сложные эфиры дикарбоновой хлорофиллиновой кислоты с двумя спиртами (фитолом и метанолом). В карбоксильных группах хлорофиллиновой кислоты водород замещен остатками метилового и фитолового спиртов. Наличие в порфириновом ядре хлорофилла коньюгированной по кругу системы десяти двойных связей и магния обуславливает характерный для хлорофилла зеленый цвет. Хлорофиллу а присущ темно-зеленый цвет, а хлорофиллу b - светло-зеленый цвет. Остаток фитола придает хлорофиллу липоидные свойства, то есть он может растворяться в жировых растворителях.
Хлорофиллам свойственна флуоресценция - т.е. свойство под влиянием падающего света, в свою очередь, излучать свет, при этом длина волны излучаемого света обычно больше длины волны возбуждающего света. В проходящих лучах цвет хлорофилла - изумрудно-зеленый, а в лучах отраженного света хлорофилл приобретает красный цвет, то есть длина волны, отражаемой хлорофиллом, больше, чем длина волны света, возбуждающего излучение хлорофилла.
Хлорофиллы различаются по спектрам поглощения, при этом у хлорофилла b по сравнению с хлорофиллом а полоса поглощения в красной области спектра несколько смещена в сторону коротковолновых лучей, а в сине-фиолетовой области максимум поглощения смещен в сторону длинноволновых (красных) лучей.
В хлоропластах листьев хлорофиллов в три раза больше, чем каротиноидов, а в плодах, лепестках, зернах, корнеплодах - наоборот.
Каротиноиды являются непременными спутниками хлорофиллов. Они подразделяются на бескислородные (каротины и ликопины, имеющие оранжевую и красную окраску - общая формула - С40Н56) и окисленные (ксантофиллы - общая формула - С40Н56О2).
Световая фаза фотосинтеза.
Световая фаза фотосинтеза проходит непосредственно в хлоропластах и состоит из поглощения пигментами хлоропластов фотонов, несущих световую энергию и превращения этой физической энергии солнца в химическую энергию макроэргических молекул. Процесс идет паралелльно по двум механизмам:
циклическому фосфорилированию в фотосистеме 1,нециклическому фосфорилированию в фотосистеме П.
При этом фотосинтетическом фосфорилировании физическая суть процесса состоит в поглощении молекулой пигмента кванта света, переход электрона, возбужденного этим квантом света, на более высокий уровень на период 10-9 -10-8 секунды, после чего электрон возвращается на прежний энергетический уровень, а поглощенная энергия затрачивается на присоединение фосфатной группы к АДФ и образование АТФ, а также на фотолиз воды и образование НАДФ. Н2.
В фотосистемах пигменты образуют реакционные центры, куда входят молекулы хлорофиллов, каротиноидов и ферментов. В каждый реакционный центр входят 2 молекулы хлорофилла, две молекулы феофитина, молекула цитохрома, ферредоксина и НАДФ. Несколько реакционных центров объединяются в фотосинтетическую единицу - комплекс пигментов и других молекул. Одна фотосинтетическая единица включает до 300 молекул хлорофилла и 50 молекул каротиноидов.
Один реакционный центр способен поглотить 50 квантов солнечного света за 1 секунду. Поглощение квантов света происходит последовательно каждой из молекул пигмента (один раз в 0,1 секунды). Синглетное состояние электронов (возбужденное состояние, при котором происходит переход электрона на более высокий энергетический уровень) длится всего около 10-9секунды, если же поступление энергии происходит стабильно, то возникает метастабильное или триплетное состояние электрона, которое длится уже 10-2секунды, которое и играет в световой фазе фотосинтеза определяющую роль.
Фотосистема 1.
Фотосистема 1, где происходит циклическое фосфорилирование, эволюционно более ранняя, процесс идет без выделения кислорода. Основой является комплекс пигментов, воспринимающие длину волны солнечного света 700 нм, при этом происходит возбуждение молекулы и образование молекул АТФ. При возбуждении электрона в фотосистеме 1 происходит его захват на более высоком энергетическом уровне белком ферредоксином, обратный путь электрона проходит с помощью цитохромов и флавопротеидов. Процесс перехода по цепи указанных ферментов определяет высвобождение энергии и передачу ее в процесс фосфорилирования АДФ.
Фотосистема П.
В процессе эволюции у высших растений сформировалась дополнительная фотосистема - фотосистема П - которая стала наиболее существенной в процессе фотосинтеза высших растений. Основой действия фотосистемы П является комплекс пигментов, воспринимающих длину волны солнечного света 680 нм. Эти пигменты образуют реакционный центр, в котором помимо реакции циклического фосфорилирования и образования молекул АТФ происходит и разложение молекулы воды и образование молекул НАДФ восстановленное, то есть нециклическое фосфорилирование. При этом активированные электроны передаются по цепи ферментов, в том числе и ферредоксинов на молекулу НАДФ, превращаясь в НАДФхН2.
В фотосистеме П на уровень переданного электрона поступает электрон от гидроксила (продукта ионизации воды), при этом два иона гидроксила соединяются и образуют перекись, которая затем разлагается, что приводит к выделению молекулярного кислорода. Важными кофакторами процессов фотосинтеза являются ионы Мn и Сl.
Фотолиз воды или реакция Хилла.
Фотолиз воды при фотосинтезе смоделирован Хиллом, поэтому предложенную им реакцию используют в качестве чувствительного фотосинтетического параметра при характеристике световой фазы фотосинтеза. В общем виде реакция выглядит так:
Похожие работы

... , в которых выращивают растения и проводят опыты в условиях определенного состава воздуха, нужной температуры и освещения. Применяя эти методы, физиологи исследуют растения на молекулярном, субклеточном, клеточном и организменном (интактное растение) уровнях. Сейчас в биологических исследованиях широко применяют электронные микроскопы просвечивающего типа с разрешающей способностью 0,15—0,5 нм, в ...
... или транспирация представляют совокупность химических и физических процессов. Чтобы понять механизм физиологического процесса, необходимо выделить его физические и химические компоненты. В связи с этим физиология растений все более нуждается в усовершенствованных методах биохимии. Биохимический подход оказался очень плодотворным в исследованиях таких сложных процессов, как фотосинтез и дыхание. ...
... насекомое тем временем усердно собирает сладкий нектар, предложенный цветком в награду за труды. Цветки правильной формы, то есть имеющие насколько осей симметрии, называются актиноморфными. В классе двудольных растений наблюдается богатое разнообразие цветковых форм. 3. Описание некоторых семейств класса двудольных Семейство лютиковых объединяет около 1500 видов трав и кустарников, многие ...
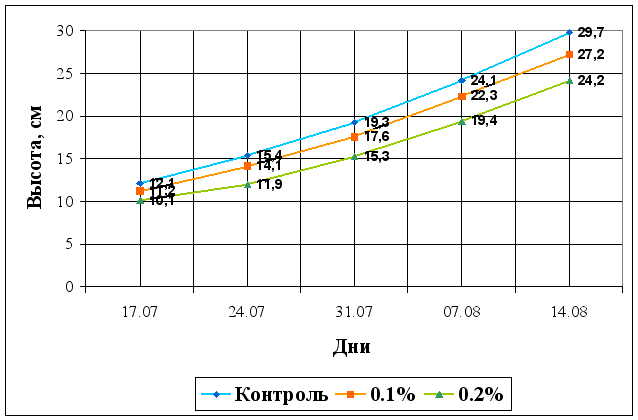
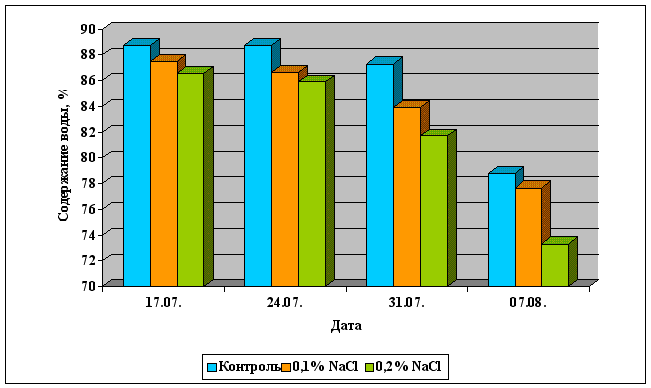
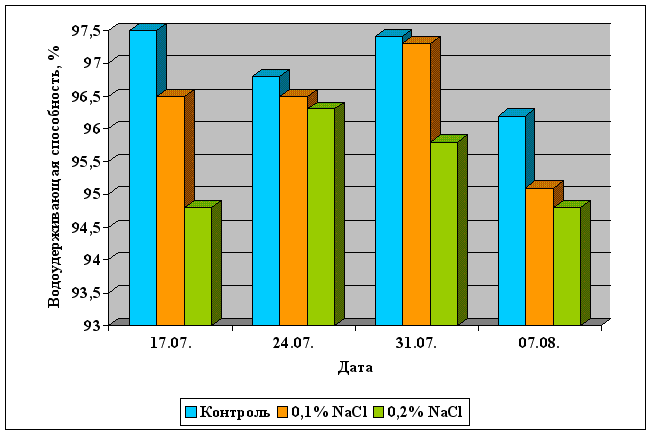
... Областного Университета в период с июня по сентябрь 2003 года. Объектом изучения являлись растения кукурузы сорта Россо. Изучение влияния 6-БАП на рост и биометрические показатели растений кукурузы при разном уровне засоления проводилось в условиях вегетационного опыта (почвенные культуры). Семена кукурузы проращивались при температуре 20оС в термостате в течение 3 дней, а затем высаживались в ...
0 комментариев