Навигация
1.1.2 Нуклеозиды и нуклеотиды
ДНК и РНК состоят из мономерных единиц - нуклеотидов, поэтому нуклеиновые кислоты называют полинуклеотидами.
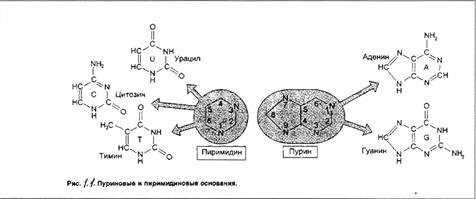
Строение нуклеотидов. Каждый нуклеотид содержит 3 химически различных компонента: гетероциклическое азотистое основание, моносахарид (пентозу) и остаток фосфорной кислоты. В состав нуклеиновых кислот входят азотистые основания двух типов: пуриновые - аденин (А), гуанин (G) и пиримидиновые - цитозин (С), тимин (Т) и урацил (U) ~ см. рисунок 1.1.
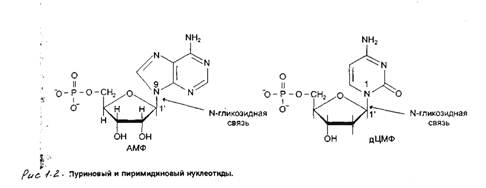
Пентозы в нуклеотидах представлены либо рибозой (в составе РНК), либо дезоксирибозой (в составе ДНК). Чтобы отличить номера атомов в пентозах от нумерации атомов в основаниях, к цифре добавляют штрих (') – 1’, 2', 3', 4' и 5'. Пентозу соединяет с основанием N-гликозидная связь, образованная Сг атомом пентозы (рибозы или дезоксирибозы) и N1-атомом пиримидина или N9- атомом пурина.
Нуклеозиды - соединения, состоящие из остатков азотистого основания и углевода - рибозы или дезоксирибозы.
Нуклеотид - это нуклеозид, к которому присоединены молекулы фосфата - см. рисунок 1.2.
Остов нуклеиновой кислоты имеет одинаковое строение по всей длине молекулы и состоит из чередующихся групп - пентоза - фосфат - пентоза. Уникальность структуры и функциональная индивидуальность молекул ДНК и РНК определяется их первичной структурой - последовательностью азотистых оснований в полинуклеотидной цепи /4/.
1.1.3 Первичная структура ДНК
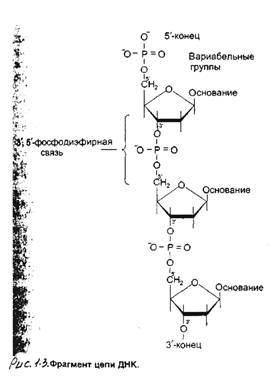
Первичная структура ДНК - порядок чередования дезоксирибонуклеозидмонофосфатов в полинуклеотидной цепи. Каждая фосфатная группа в полинуклеотидной цепи, за исключением фосфорного остатка на 5 '-конце молекулы, участвует в образовании двух эфирных связей с участием 3' и 5 '-углеводных атомов двух соседних дезоксирибоз, поэтому связь между мономерами обозначают 3',5' -фосфодиэфирной.
Концевые нуклеотиды ДНК различают по структуре: на 5'-конце находится фосфатная группа, а на 3'-конце цепи - свободная ОН-группа. Эти концы называют 5' и 3'-концами — см рисунок 1.3.
Линейная последовательность дезоксирибонуклеотидов в полимерной цепи ДНК обычно сокращенно записывают с помощью однобуквенного кода, например A-G-C - Т- Т- А- С- А от 5'-к З '-концу.
В каждом мономере нуклеиновой кислоты присутствует остаток фосфорной кислоты. При рН 7 фосфатная группа полностью ионизирована, поэтому in vivo нуклеиновые кислоты существуют в виде полианионов (имеют множественные отрицательный заряд). Остатки пентоз тоже проявляют гидрофильные свойства. Азотистые основания почти нерастворимы в воде, но некоторые атомы пуринового и пиримидинового циклов способны образовывать водородные связи.
1.1.4 Вторичная структура ДНК
Молекула ДНК имеет форму спирали, образованную двумя полинуклеотидными цепями, закрученными относительно друг друга и вокруг общей оси. Двойная спираль правозакрученная, полинуклеотидные цепи в ней антипараллельны, т.е. если одна из них ориентирована в направлении 3' —> 5', то вторая в направлении 5'—> 3'. Поэтому на каждом из концов молекулы ДНК расположены 5' -конец одной цепи и 3'-конец другой цепи.
Двойная спираль стала символом новой, молекулярной биологии. Спираль построена за счёт комплементарных оснований. Именно двухцепочечностью и комплементарностью оснований объясняются биологические функции ДНК. В правовинтовой спирали две водородные связи в паре А - Т и три водородные связи в паре G - С . Естественно, последняя пара прочнее, а первая слабее. Модель Уотсона Крика объяснила правило Чаргаффа - большое основание - пуриновое дополняется маленьким основанием - одиночным гетероциклом пиримидина. То же самое вG-CивA-T. Благодаря этому размеры пар одинаковы по ширине, что и приводит к регулярной двойной спирали. Спираль не деформируется, поскольку то, что её скрепляет внутри, симметрично по отношению друг к другу, хотя и повёрнуто. Плоскости азотистых оснований наклонены по отношению к оси спирали образуя третичную структуру. В местах сгиба двойная спираль имеет изломы, то есть нарушается классическая двойная структура ДНК.
1.2 Эпигенетические механизмы наследственности
1.2.1 Контроль активации генов
Существует мнение, что все свойства клетки и даже многоклеточного организма однозначно определены последовательностью нуклеотидов в ДНК. В настоящее время это положение подвергается критике. Выдвинуты представления об эпигенетических механизмах избирательной экспрессии генов, ведущая роль в которых принадлежит хроматину. Именно на уровне хроматина работают основные механизмы программирования развития живого организма. Эти механизмы базируются на структурно-функциональных особенностях нуклеосомы, гистонов, формирующих нуклеосомную сердцевину, межнуклеосомных взаимодействиях, тонко регулируемых внешними сигналами. На уровне нуклеосомы реализуется «гистоновый код», управляющий сложнейшим каскадом биохимических реакций в клетке. Все вышеуказанные механизмы обеспечивают важнейшее приспособление организмов к изменяющимся условиям внешней и внутренней среды.
Геном каждой клетки человека составляет около 40 тысяч генов, но все они одновременно не используются. В каждом клеточном типе работает около 20 тысяч генов. Как минимум половина всех работающих генов необходима для поддержания жизнедеятельности любой клетки, другая половина определяет специализацию клетки, т.е. будет ли она принадлежать печени, почкам, селезёнке, другому органу или ткани. Все клетки содержат одинаковую информацию, записанную в молекуле ДНК, но при развитии организма эта информация в тканях или органах считывается избирательно, что и приводит к огромному разнообразию клеток в организме.
Существует несколько уровней контроля активации генов. Один из них - модификация ДНК, соответствующая тем генам, которые необходимо выключить, т.е. этот ген должен молчать в данном типе клеток. При этом данный признак сохраняется и при удвоении клетки. Однако ДНК сама по себе несёт важнейшую функцию кодирования аминокислотной последовательности белков, и человек не может произвольно её модифицировать, не затронув этой информации и не изменив способность ДНК к удвоению.
На сегодня единственной изученной модификацией ДНК, которая не нарушает кодирование и копирование, является метилирование цитозина, которое выключает ген и он становится молчащим, и, главное, это свойство закрепляется в наследственности, т.е. в дочерней клетке сохраняется информация о том, что данный ген должен молчать.
Как же эта модификация влияет на активацию гена - на получение РНК- копии данного гена, служащей в дальнейшем матрицей для синтеза белка?
Указанная модификация копируется при удвоении ДНК. Возможны два механизма контроля активации гена. Первый - отталкивание белков, активирующих ген, второй - привлечение к метилированной ДНК белков, которые селективно узнают только метилированную ДНК и участвуют в очень плотной её упаковке за счёт изменения структуры хроматина - формы существования генома высших организмов. Именно на уровне хроматина происходит основной контроль активации генов.
Благодаря расшифровке генома возникли две новые науки: геномика, изучающая геномы, и протеомика, изучающая совокупность клеточных белков. Почти 50 лет после открытия ДНК господствовал постулат: один ген синтезирует один белок. После расшифровки генома появилась новая догма: один ген - много белков. Установлено, что гены у высших организмов состоят из кодирующих и некодирующих участков. В результате комбинаторики из одного гена путём соединения кодирующих участков можно получить очень много белков. Если у человека около 40 тысячи генов (по геномике), то по протеомике можно получить минимально более 100 тысяч белков. Это только на уровне первичной структуры, фактическая ситуация ещё сложнее, потому что белки подвергаются модификациям, на них «навешивается» огромное количество разных групп, что приводит к колоссальному разнообразию. Если геном постоянен, то протеом очень вариабелен. Уотсон очень много сделал для расшифровки генома человека, которая закончилась в 2003 году /3/.
0 комментариев