Навигация
Денатурирующий градиентный гель-электрофорез
37420
знаков
0
таблиц
7
изображений
1.2 Денатурирующий градиентный гель-электрофорез
Два фрагмента ДНК, различающиеся лишь делецией, инсерцией или заменой одного нуклеотида, или единственным неспаренным нуклеотидом, можно легко разделить при помощи денатурирующего градиентного гель-электрофореза, ДГГЭ. Принцип метода заключается в разделении двухцепочечных фрагментов ДНК электрофорезом в стандартном акриламидном геле с линейным градиентом денатурирующих факторов – мочевины, формамида или температуры. Ниже приводятся теоретические основы метода. Здесь отметим только, что разделение очень похожих фрагментов в геле происходит из-за разных температур плавления их ДНК.
1.2.1 Теоретические основы метода ДГГЭ
При постепенном повышении температуры или концентрации денатурирующего вещества происходит плавление двухцепочечной ДНК, и для каждого фрагмента параметры этого процесса различны. Внутри фрагмента имеются так называемые области плавления – блоки последовательностей, плавящихся одновременно при определенных дискретных значениях температуры, которая и является температурой плавления данной области. Длина таких областей составляет от 25 до нескольких сотен нуклеотидных пар. Два соседних домена могут иметь четкую границу и различаться по Гт на несколько градусов. Фрагменты ДНК длиной 100–1000 п.н. имеют обычно от двух до пяти областей плавления. Температура плавления доменов в рестрикционных фрагментах большинства образцов ДНК из природных источников колеблется от 65 до 80°С.
Общеизвестно, что нуклеотидный состав фрагментов ДНК влияет на температуру их плавления. Оптимальные условия для отжига и отмывки нуклеиновых кислот в реакциях гибридизации подбирают исходя из их нуклеотидного состава. В гораздо меньшей степени учитывается вклад стэкинг-взаимодействий в термодинамическую стабильность двойной спирали. А между тем энергия таких взаимодействий между соседними нуклеотидами одной цепи ДНК, удерживающих ее в скрученном состоянии в физиологических растворах, больше энергии водородных связей комплементарного спаривания. Порядок чередования оснований определяет степень стэкинга и, следовательно, влияет на термостабильность фрагмента ДНК. Даже единичная нуклеотидная замена может так сильно сказаться на стэкинг-взаимодействий, что Гт изменится более чем на 1°С. Поскольку процесс плавления домена практически полностью определяется кооперативными взаимодействиями, любая единичная нуклеотидная замена в любой его точке будет менять температуру плавления,
Разработанный Лерманом и Фишером метод электрофоретического разделения близких последовательностей ДНК основан на использовании различий в температуре плавления, обусловленных нуклеотидными заменами. Они исходят из того факта, что последовательность оснований влияет на Тт области плавления и что конформация фрагмента ДНК обусловливает его подвижность в геле под действием электрического поля. Принцип метода – электрофорез ДНК в акриламидном геле фиксированной концентрации с линейно возрастающим к нижней части геля градиентом концентрации веществ, денатурирующих ДНК: обычно мочевины или формамида. Электрофорезная камера нагревается до температуры примерно 60°С.
Это чуть ниже температуры плавления самой легкоплавкой области во фрагменте ДНК – двухцепочечная молекула ДНК входит в гель и продвигается в нем со скоростью, пропорциональной ее молекулярной массе. Как только фрагмент достигает участка геля, в котором температура камеры и концентрация денатурирующих веществ создают условия для плавления первой, самой легкоплавкой области, он как бы разветвляется: одна часть его остается двухцепочечной, а другая переходит в одноцепочечную форму. Такая разветвленная структура застревает в порах геля, в результате чего ее подвижность снижается. Снижение электрофоретической подвижности молекулы коррелирует с соотношением длин денатурированного и двухцепочечного участков: чем длиннее самый легкоплавкий домен, тем сильнее снижается подвижность. Участок геля, в котором молекула ДНК начинает терять скорость, соответствует самым легкоплавким доменам. Если концентрации денатурирующих веществ подобраны правильно, то два фрагмента ДНК, различающиеся лишь одной нуклеотидной заменой и имеющие неодинаковые значения Тт., начнут снижать скорость в различных участках геля, и к концу прохождения через него их легко будет разделить. Ранее считалось, что отделить мутировавшие фрагменты от фрагмента ДНК дикого типа за счет разной степени снижения их подвижности в ДГГЭ можно лишь в том случае, если замены произошли в области с самым низким значением Тт. Однако анализ большого числа образцов ДНК, а также последующие теоретические выкладки показали, что в большинстве случаев метод ДГГЭ дает возможность обнаружить нуклеотидные замены во всех областях плавления, за исключением самых термостабильных. Замены в областях с самой высокой температурой плавления обычно не обнаруживаются, так как по окончании плавления последнего домена ДНК полностью денатурирует, а это приводит к нарушению корреляции между конформацией молекулы и скоростью ее продвижения. Важно помнить, что успех при проведении ДГГЭ с целью разделения мутировавшего фрагмента и фрагмента ДНК дикого типа определяется структурой фрагмента. Она должна быть разветвленной, причем мутации должны приходиться на области, подвергшиеся плавлению. Для увеличения разрешающей способности метода к исследуемому фрагменту ДНК «пришивали» последовательность с более высокой температурой плавления ДНК, так называемый «GC-зажим». При этом все области плавления фрагмента, включая самую тугоплавкую, становились доступными для анализа. Эти опыты проводили с препаратами ДНК, клонированными в плазмидном векторе, содержащем «GC-зажим». Чтобы избежать стадии клонирования, фрагменты с «зажимами» можно получить в полимеразной цепной реакции.
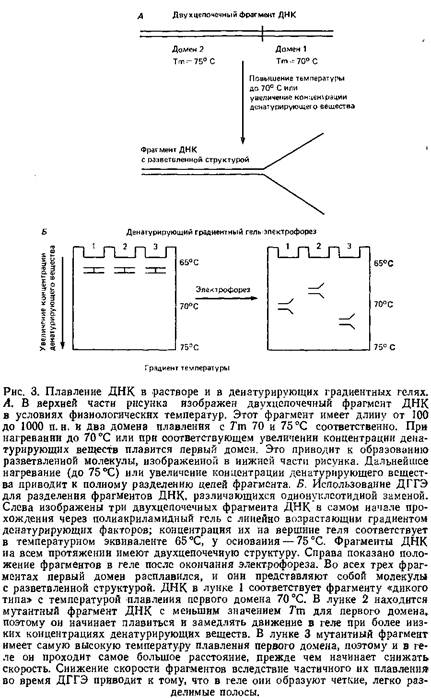
Еще одним усовершенствованием ДГГЭ следует считать использование гетеродуплексных фрагментов, образованных мутировавшей ДНК и ДНК дикого типа. В предыдущих опытах сравнивали подвижность гомодуплексных фрагментов мутантной и исходной ДНК в параллельных дорожках при ДГГЭ. Работа с полученными из них гетеродуплексами позволяет значительно повысить разрешающую способность метода и долю обнаруживаемых мутаций. Причина заключена в том, что неспаренные участки гетеродуплекса, в которых имеются однонуклеотидные замены, сильно снижают стзкинг-взаимодействия оснований, дестабилизируя вторичную структуру, а это, в свою очередь, приводит к снижению температуры плавления фрагмента ДНК- В ряде случаев замена всего одного нуклеотида обусловливала снижение температуры на целых 6°С. Анализ большого числа гетеродуплексов и теоретические подсчеты показывают, что все возможные замены нуклеотидов в легкоплавких доменах, приводящие к образованию неспаренных участков во фрагменте ДНК, выявляются при помощи ДГГЭ с вероятностью 100% за счет изменения подвижности. На долю легкоплавких областей во фрагментах ДНК от 100 до 1000 п.н. приходится в среднем более 50% их длины, следовательно, использование гетеродуплексов таких фрагментов позволяет выявить более половины всех возможных замен нуклеотидов. В клонированных же фрагментах ДНК с «GC-зажимом» выявляются практически все замены нуклеотидов.
Гетеродуплексы дают возможность наблюдать изменение подвижности мутантных фрагментов ДНК в геле, не выявляемое при стандартных условиях проведения электрофореза. Зачастую единичные мутации можно обнаружить по вызываемым ими значительным изменениям структуры доменов во фрагменте ДНК-Изменения эти отражаются и на характере плавления ДНК: домены, имевшие изначально самую высокую температуру плавления, могут стать самыми легкоплавкими. Таким образом, когда вместо гомодуплексов используются гетеродуплексы, ДГГЭ позволяет выявлять замены нуклеотидов даже в самых тугоплавких доменах.
Похожие работы
... , вызванные динамическими му-тациями.-----------------------T-----------T-------T-----T------T------T----------------------¬ Болезнь, номер по ¦ Ген, лока-¦Триплет¦Норма¦Прему-¦Мута- ¦Литература ¦ МакКьюсику (MIM) ¦ лизация ¦ ¦ ¦тация ¦ция ¦ ¦ -----------------------+-----------+-------+-----+------+------+----------------------+ Синдром ломкой X-хро- ¦FMR1, FRAXA¦(CGG)n ...
0 комментариев