Навигация
Контроль афферентного входа в соматосенсорной системе
85062
знака
0
таблиц
15
изображений
2.12 Контроль афферентного входа в соматосенсорной системе
На всех уровнях ЦНС афферентная информация может испытывать различные влияния и изменяться (модулироваться). Это вызывается либо взаимным торможением афферентами друг друга (афферентным торможением), либо центробежным (нисходящим) торможением. Несколько примеров афферентного торможения уже рассматривались: это аутогенное и антагонистическое торможение в спинальных двигательных системах и латеральное торможение во многих сенсорных системах.
Центробежная регуляции афферентных сигналов
Сенсорные системы нельзя рассматривать как улицы с односторонним движением, по которым информация передается от периферии к коре; напротив, на промежуточную переработку такой информации влияет целый ряд (возбуждающих или тормозных) сигналов центрального происхождения, проводимых в противоположном направлении. Схема на рис. 2.7, А показывает нисходящие тормозные пути соматосенсорной системы, начинающиеся в коре и стволе мозга.

Рис. 2.7. Центробежная регуляция соматосенсорной системы. А. Афферентная информация может модулироваться в синапсах нисходящим торможением. Б. Пример функций нисходящего торможения. В синапсах, через которые активность афферентов (а, б, в) передается центральному соматосенсорному нейрону, оно может изменять величину рецептивного поля этого нейрона, если афференты, идущие от периферической части рецептивного поля (а, в), тормозятся сильнее, чем идущие от его центра (б)
Что обеспечивают эти системы нисходящего торможения? Их известные или предполагаемые функциональные влияния на афферентную информацию можно обобщить следующим образом:
- порог афферентной синаптической передачи повышается. Одно из следствий этого – подавление обыденной информации (например, непрерывной стимуляции одеждой);
- размеры рецептивного поля центрального нейрона уменьшаются при усилении нисходящего торможения (рис. 2.7, Б);
- изменяется модальность нейрона, на котором конвергируют разные типы афферентов (рис. 2.7, Б, а, б);
- регулируется чувствительность или устанавливаются пределы передачи афферентной информации.
Регуляция чувствительности афферентной передачи нисходящим торможением

Рис. 2.8. Торможение соматосенсорной информации в спинном мозгу стимуляцией среднего мозга. А. Схема нисходящей системы и экспериментальной методики ее изучения. Стимулирующий электрод вживлен в центральное серое вещество (ЦСВ) среднего мозга наркотизированного животного. Другой электрод регистрирует здесь активность одиночных нейронов в ответ, например, на раздражение кожи очень сильным нагреванием. Б. Peaкция спинального нейрона на нагревание кожи до (слева) и во время (справа) электрической стимуляции ЦСВ. Зависимость между частотой импульсации спинального нейрона (ось ординат) и интенсивностью нагревав (температура кожи, ось абсцисс) до и во время стимуляции ЦСВ.
Рис. 2.8 показывает, что афферентная передача от кожных рецепторов может тормозиться в нейронах спинного мозга электрической стимуляцией среднего мозга. Тормозные стимулы идут по нисходящему пути и действуют либо пре-, либо постсинаптически. Тормозятся как ноцицептивные, так и неноцицептивные сигналы. В этом примере сенсорным стимулом служит температура кожи повреждающих уровней; ее уровень кодируется приблизительно линейно частотой импульсации спинальных нейронов (рис. 2.8, В). Стимуляция среднего мозга ее снижает (рис. 2.8, Б), из-за чего наклон кривой, отражающей кодирование интенсивности, уменьшается (рис. 2.8, В). Здесь нисходящее торможение можно рассматривать как механизм настройки чувствительности или изменения коэффициента усиления афферентной передачи; реакции на стимулы при любой интенсивности последних уменьшаются в одинаковое число раз в зависимости от степени нисходящего торможения.
О том, каким образом такие тормозные системы активируются в естественных условиях, известно еще мало. Одна из их функций, как полагают,- содействовать вниманию. Было высказано предположение, что иногда информация, восходящая от спинного мозга, может сама активировать нисходящее торможение. Это был бы случай возвратного торможения (торможения по обратной связи), отличающегося от спинального сегментарного возвратного (через клетки Реншоу) торможения с участием супраспинальной петли. В этом особом cлучае торможения по обратной связи его эффект может выражаться в автоматической установке пределов: наклон характеристической кривой (рис. 2.8, В), т. е. коэффицент усиления афферентной передачи, определяется интенсивностью ее самой. Такой механизм сравним, например с автоматической регуляцией коэффициента усиления во входе кассетного магнитофона.
Влияние двигательных систем на афферентную информацию
Двигательные центры тоже могут оказывать значительное центробежное влияние на сигналы, поступающие от рецепторов. Возьмем контроль состояния мышечного веретена системой ![]() -волокон, регуляцию положения глаз глазодвигательной системой, ощупывающие движения пальцев при тактильном исследовании и изменения мышечного тонуса в аппарате среднего уха. Их считают механизмами центробежной модификации сенсорных каналов. Эти примеры ясно показывают, что ЦНС участвует в восприятии, не только пассивно принимая периферическую информацию, но и активно влияя на поток информации и регулируя его множеством способов.
-волокон, регуляцию положения глаз глазодвигательной системой, ощупывающие движения пальцев при тактильном исследовании и изменения мышечного тонуса в аппарате среднего уха. Их считают механизмами центробежной модификации сенсорных каналов. Эти примеры ясно показывают, что ЦНС участвует в восприятии, не только пассивно принимая периферическую информацию, но и активно влияя на поток информации и регулируя его множеством способов.
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
1. Константинов А.И., Соколов В.А., Быков К.А. Сравнительная физиологическая сенсорная система. – М., 1980.
2. Серопегин И.М., Волков В.М., Синайский М.М. Физиология человека. – М., 1979.
3. Измайлов Ч.А. Психофизиология цветового зрения. – М., 1989.
4. Бертулис А.В., Глезер В.Д. Провтранственное цветовое зрение. – Л., 1990.
5. Шевелев И.А. Динамика зрительного сенсорного сигнала. – М., 1971.
6. Марр Д. Зрение. – М., 1987.
7. Дудкин К.Н. Зрительное восприятие и память. – Л., 1985.
8. Гуртовой Г.К. Глаз и зрение. – М., 1959.
9. Барер А.С., Сокол Е.А. Функциональное состояние зрительного анализатора человека – оператора. – М., 1990.
10. Зимкина Н.В. Физиология человека. – М., 1975.
11. Васильева В., Коссовская Э., Степочкина Н. Физиология человека. – М., 1973.
12. Физиология человека. / Под. Ред. Шмидта Р., Тевса Г. – Т1, М., 1996.
13. Общий курс физиологии человека и животных. – Т.1, М., 1991.
14. Основы физиологии человека. – Т.1, М., 1994.
Похожие работы
... у человека. Длина волны, одинаково возбуждающая оба типа колбочек (в области пересечения кривых поглощения), воспринимается как белый цвет и называется «нейтральной точкой» спектра. Такое смешение меньше выражено в зрительных системах с тремя типами цветовых рецепторов или при трихроматическом зрении), известном у многих видов, в том числе у человека. Однако некоторое смешение происходит и здесь: ...
... состава хрусталика и стекловидного тела, ведущие к их помутнению. Поэтому при преломлении света наблюдается его диффузное рассеивание. При рассмотрении белого фона поверхности человек видит мелькающие кружочки, точки и т. д. 2.4 Зрительное восприятие 2.4.1 Поля зрения Если фиксировать глазом какую-либо точку, ее изображение падает на желтое пятно. И в этом случае мы видим точку ...
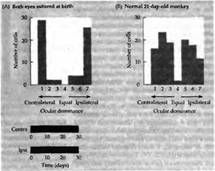
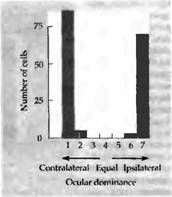
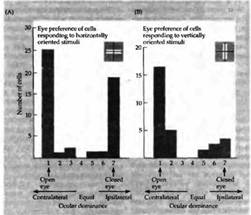
... депривированному глазу. Подобные же результаты были получены Бонхоеффером с коллегами на котятах, которым представлялись только полоски определенной ориентации. Критические периоды в развитии зрительной системы человека и их клиническое значение Чувствительность котят и детенышей обезьян в ранние периоды жизни напоминает клинические наблюдения, выполненные на людях. Давно известно, что ...
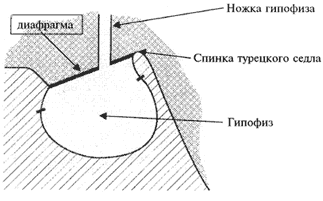
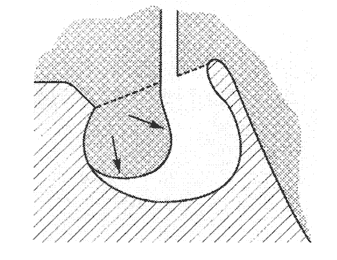
... нарушений. Как правило, болезнь протекает благоприятно при ранней диагностике и адекватной симптоматической терапии. Тяжелые прогрессирующие зрительные нарушения являются показаниями для хирургического лечения. Список литературы 1. Бабарина М.Б. Клинико–гормональные аспекты синдрома «пустого» турецкого седла: Автореф. дис. канд. мед. наук. – М., 1999. 2. Вегетативные расстройства /Под ред ...
0 комментариев