Фактори впливу у водному середовищі
Гідробіоценози як біологічні системи гідросфери
Структура гідробіоценозів
Сапробність водних екосистем
Токсичне забруднення та його наслідки для водних екосистем
Екологічний стан водойм України
Результати моніторингу Кременчуцького водосховища
Екологічна оцінка стану якості Кременчуцького фодосховища
Навигация
Структура гідробіоценозів
Гідробіоценоз Кременчуцького водосховища
78254
знака
2
таблицы
5
изображений
1.2.4 Структура гідробіоценозів
Гідробіоценози, які складаються з автотрофних і гетеротрофних організмів, називаються повночленними. Інколи можуть зустрічатись водні екосистеми, в яких практично відсутні автотрофні організми, а є лише гетеротрофні. До таких неповночленних гідробіоценозів належать біоценози водойм темних печер, де не може протікати фотосинтез. У той же час у них можуть жити найпростіші, хемосинтезуючі бактерії і навіть деякі безхребетні, які живляться готовими органічними речовинами, що надходять з інших джерел (наприклад, з фільтраційних вод, які проходять через шар грунту і вимивають з нього не тільки неорганічні, а й органічні речовини).
До складу повночленних гідробіоценозів входять гідробіонти різних систематичних груп: мікро- та макроводорості, вищі водяні рослини, бактерії, актиноміцети, найпростіші, безхребетні і риби [2].
Гідробіоценози можна називати і характеризувати як угруповання водоростей (альгоценози), вищих водяних рослин (фітоценози), тварин (зооценози), риб (іхтіоценози) тощо. Можна виділяти гідробіоценози також за характером біотопу, наприклад, гідробіоценози товщі води, літоральні зони, скель (морські біоценози), піщаного грунту тощо. В гідроекологічній практиці нерідко об’єктом системного дослідження бувають не всі популяції гідробіоценозу, а тільки ті, що належать до певного таксону (таксоценоз).
Залежно від характеру донних грунтів може змінюватись і донна фауна. За цією ознакою в континентальних водоймах розрізняють біоценози піщаних грунтів (псамофільні), глинистих грунтів (аргілофільні), кам’янистого дна (ліофільні), мулу (пелофільні). Для проточних (лотичних) водних систем до назви біоценозу додається префікс “рео”. Так, біоценози річкових систем з піщаним дном мають назву псамореофільні, з глинистими — аргілореофільні, з мулистими — пелореофільні тощо.
Для водного середовища характерною є вертикальна структура гідробіоценозів. Вона визначається, перш за все, екологічними умовами, характерними для окремих екологічних зон водних об’єктів. Так, для пелагічних зон визначальними факторами є градієнт освітленості, температура, газовий режим, концентрація біогенних речовин. На великих глибинах морів і океанів формування донних біоценозів залежить від гідростатичного тиску, характеру грунтів та динаміки водних мас. Усі ці чинники визначають специфіку видового складу, переважання певних видів, їх біопродуктивність та вплив на популяції інших організмів. Саме абіотичні фактори середовища визначають просторову структуру гідробіоценозів. Просторовим межам гідробіоценозів відповідає біотоп, для якого мінімальний простір з комплексом взаємодіючих видів забезпечує повний цикл біогенного кругообігу речовин і енергії [4].
Угруповання вищих водяних рослин (фітоценози) характеризуються як горизонтальною, так і вертикальною структурою. Окремі ділянки заростей можуть відрізнятися щільністю, віковим складом та загальною фітомасою. Просторовий поділ пов’язаний з підтриманням оптимальної щільності рослин в умовах ценотичної конкуренції.
Вертикальна структура фітоценозів формується у відповідності з рівнем освітленості, який впливає на фотосинтетичну активність популяцій в цілому. Її можна характеризувати, виходячи з ярусності рослинності, тобто співіснування різних за висотою груп рослин в межах фітоценозу. В межах фітоценозу можна спостерігати дво- і триярусну структуру, яка утворюється популяціями інших видів рослин. Серед вільноплаваючих вищих водяних рослин виділяються масовим поширенням фітоценози сальвінії плаваючої, яка в місцях масового розвитку вкриває поверхню водойм суцільним килимом. З прикріплених рослин з плаваючими вегетативними органами, які можуть утворювати до 100% проектного покриття, можна вказати фітоценози латаття білого. Вони часто мають двоярусну вертикальну структуру [5].
Таким чином, форми структурної організації фітоценозів водяних рослин досить різноманітні. Поряд з невеликими куртинами вони можуть утворювати суцільні зарості, які займають значні площі мілководь. При цьому вертикальна ярусність у них пов’язана з генетично закріпленою особливістю протікання всіх фізіологічних процесів (фотосинтез, утилізація поживних речовин, транспірація, вегетативне та статеве розмноження).
Структура планктонних і донних ценозів найбільш наглядно демонструється графічно у вигляді ценограми, яка поєднує два компоненти: графік, що відображає ранжування видових популяцій за рівнем їх кількісного розвитку, і секторіальну діаграму, на якій показані відсоткові співвідношення (за біомасою, чисельністю) основних таксономічних груп (для фітопланктону — зелені, діатомові, синьо-зелені та інші водорості; для зоопланктону — гілястовусі та веслоногі ракоподібні, коловертки; для зообентосу — молюски, олігохети, личинки хірономід та інших комах тощо). У центрі малого кола, вписаного у велике, вказується загальна біомаса ценозу [3].
1.2.5 Взаємини гідробіонтів в екосистемах
Гідробіонти в екосистемах перебувають у постійному контакті та досить складних взаємних стосунках. Найважливіші з них — це топічні та трофічні (харчові) зв’язки.
Топічні відносини пов’язані із спільним життям різних популяцій в одному біотопі і відповідно — міжвидовою конкуренцією за місце, розчинений кисень та інші необхідні для життя умови.
Трофічні зв’язки, що тісно переплітаються, формують трофічні ланцюги різного типу, по яких циркулюють потоки енергії. Різні види мають і свої і спільні з іншими джерела живлення, а переплетіння трофічних ланцюгів створює трофічну сітку екосистеми.
Кожний член трофічного ланцюга виступає по відношенню до попередніх як хижак, а по відношенню до тих, що його споживають, — як жертва. Наприклад, гіллястовусі ракоподібні поїдають планктонні водорості і мікроорганізми, їх самих поїдають риби-планктофаги (мирні риби), а останніх — хижі риби. Паралельно існують інші трофічні ланцюги: коловертки поїдають мікроводорості і бактерії, хижі циклопи поїдають коловерток і молодь гіллястовусих, а самих циклопів — риби-планктофаги.
В.М. Беклемішев виділяє ще два типи зв’язків: міграційні, або форичні, і фабричні. Останній тип зв’язків виникає тоді, коли організми одного виду використовують виділення (екскременти) або зруйновані частини тіла іншого організму (наприклад, черепашки молюсків, скинуті шкурки злинялих ракоподібних) як місце для оселення або для побудови власного тіла.
Окремий тип міжпопуляційних зв’язків становлять антибіотичні зв’язки, які пов’язані з виділенням гідробіонтами, зокрема вищими водяними рослинами, антибіотиків, що згубно діють на мікроби, віруси та водорості, а також різних отруйних речовин (синьо-зелені, динофітові, отруйні безхребетні та риби). Виділення антибіотиків та отруйних речовин — це “хімічна зброя” гідробіонтів у боротьбі за існування [6].
Відповідні відносини між рослинами в ботаніці називаються алелопатією.
Як наслідок взаємодії всіх співугруповань гідробіонтів та абіотичних факторів середовища формується екологічна ніша популяції, тобто те місце, яке воно займає у системі біоценотичних зв’язків. Ніша — це сукупність усіх умов, необхідних для існування виду, не обмеженого часом і простором. Вона характеризує ступінь біологічної спеціалізації виду.
Особини одного і того ж виду на різних стадіях розвитку можуть займати різні екологічні ніші, наприклад, пуголовки, що живуть в воді і входять до складу водних екосистем, і дорослі жаби, які живуть в наземному середовищі, а для розмноження знову входять в воду.
Досить складними є взаємини риб і безхребетних в екосистемах. Зоопланктон, зообентос та частково зооперифітон і зоофітос споживаються рибами і становлять їх природну кормову базу. Виїдаючи зоопланктон, риби-зоопланктофаги істотно змінюють кількісні характеристики і співвідношення популяцій і отже — структуру планктонних співугруповань. Те саме стосується взаємовідносин риб-бентофагів та бентосних організмів.
Виїдання може бути суцільним або вибірковим, що залежить від співвідношення розмірів тіла (зокрема ротового отвору) риби-споживача і кормових об’єктів. Деякі зоопланктони, наприклад, Daphnia longispina, мають вирости, голки, шипи та інші морфологічні утворення, які колють ротовий отвір риб, і тому риби обминають їх. Інших зоопланктонів, що не мають відповідних морфологічних захисних елементів, вони споживають більш охоче [7].
Інтродукція риб у водоймі може істотно змінювати цю ситуацію, бо наприклад фітопланктофаг білий товстолоб масово виїдає фітопланктон, а зоопланктофаг строкатий товстолоб виступає як додатковий споживач зоопланктону.
Виїдаючи свою природну кормову базу, риби зрештою починають голодувати. Це, зокрема, спостерігається в нерестових ставках, де велика кількість мальків, що народилися одночасно, може за короткий час виїсти весь зоопланктон і загинути від голоду.
Дефіцит кормів може виникати внаслідок міжвидової конкуренції (як між різними видами риб, так і між рибами і хижими безхребетними). Про ступінь її напруженості також можна судити, досліджувати вміст шлунково-кишкового тракту риб різних видів.
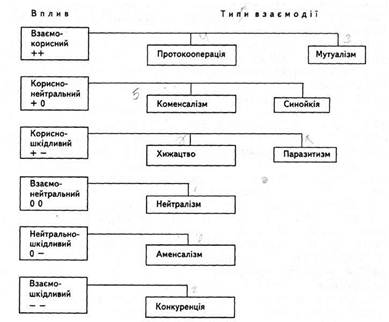
Поширеним типом зв’язків в екосистемах є симбіоз (термін “симбіоз” запропонував А. де Баріу у 1879 р.). Під ним розуміють тривале співіснування організмів різних видів, при якому обидва організми (симбіонти) краще пристосовуються до умов середовища. У симбіотичних системах можуть виникати трофічні, просторові та інші типи взаємовідносин. За характером відносин між партнерами виділяють такі основні типи симбіозу [3].
Коменсалізм є такою формою взаємовідносин між двома видами організмів, коли один з них користується якимись перевагами іншого, не завдаючи йому безпосередньої шкоди. При такій формі співіснування коменсал використовує хазяїна як місце поселення або засіб пересування, а сам виступає свого роду санітаром, який підбирає залишки його корму або живиться іншими організмами, що живуть у тілі хазяїна. Так, джгутиконосці і амеби, які живуть у кишечнику риб, живляться бактеріями, які в ньому знаходяться. Прикладом коменсалізму кишковопорожнинних і риб є постійне перебування молоді пікши, тріски та інших риб під дзвоном великої арктичної медузи Cyanea capillata. Вони використовують її як схованку та місце знаходження корму: об’їдають залишки корму, що прилипли до щупалець.
Особливою формою взаємовідносин гідробіонтів в екосистемах є мутуалізм. Це така форма співіснування, коли жоден з партнерів не може існувати без іншого. Прикладом можуть служити водорості, які живучи в організмі зеленої гідри, віддають їй до 30% продуктів фотосинтезу. Ще більше (до 50%) віддають органічних речовин інфузоріям (Paramecium caudatum) зелені водорості, які в процесі свого метаболізму засвоюють вуглекислоту з тіла інфузорій.
Мутуалізм автотрофних і гетеротрофних організмів є не винятковим явищем, а еволюційно закріпленим співіснуванням.
Однією з форм взаємовідносин між гідробіонтами в екосистемах є стимуляція. Під нею розуміють стимуляцію росту організмами одних популяцій іншими. Як правило, її основою є біохімічна взаємодія між метаболітами, які виділяються організмами, що входять до різних популяцій. Наприклад, для нормального розвитку діатомових водоростей потрібен вітамін В12, який виділяється у процесі життєдіяльності бактерій. При перебуванні у біоценозі бактерій і водоростей створюються сприятливі умови, за яких водорості виділяють достатню кількість полісахаридів, необхідних для розвитку бактерій, а останні забезпечують їх вітаміном В12. Завдяки цьому стимулюється ріст і розвиток як бактерій, так і водоростей.
Зовсім іншими взаємовідносинами характеризується конкуренція, хижацтво і паразитизм. Конкуренція — це взаємовідносини між організмами одного виду, які проявляються у боротьбі за корм, біотоп тощо. Хижацтво характеризується виживанням одних організмів за рахунок поїдання інших. Воно може набувати різних форм. Хижаками можуть бути і рослини. Так, на кільчастих листках пухирника розміщені пухирці, які мають вхідний отвір, що закривається клапаном. Оскільки листя занурене у воду, через такий отвір у пухирці заходять личинки риб і, не маючи можливості повернутись назад, вони гинуть, а продукти розкладу тіла засвоюються цією рослиною [6].
Паразитизм — це форма взаємовідносин двох різних організмів, які належать до різних видів. Він характеризується більш складним, ніж хижацтво, антагоністичним характером зв’язки. Один з видів (паразит) використовує іншого (хазяїна) як середовище перебування і джерело живлення, покладаючи на нього регуляцію своїх відносин із зовнішнім середовищем. Відносини у паразитичних системах між партнерами побудовані за принципом нестійкої рівноваги, порушення якої може призвести до загибелі одного або двох партнерів.
В залежності від належності до хазяїна, паразитів поділяють на облігатних (обов’язкових) та факультативних (не обов’язкових). У тих випадках, коли паразити нападають на хазяїв тільки під час живлення, такі взаємовідносини отримали назву тимчасовий паразитизм. За місцем розташування паразитів поділяють на ектопаразитів, що мешкають на поверхні тіла, та ендопаразитів, які живуть у внутрішніх органах (тканини, клітини, порожнини тіла).
Серед паразитів риб зустрічаються практично представники усіх класів найпростіших: джгутикових, корененіжок, споровиків та інфузорій.
У водних екосистемах паразитизм досить поширене явище, бо водне середовище сприяє передачі паразитичних організмів від одного виду тварин до інших. Відомі такі форми паразитичних взаємовідносин, коли самі паразити служать хазяями для інших паразитів. Так, деякі мікроспоридії паразитують у трематодах, цестодах та інших паразитах. Деякі з них викликають загибель акваріумних молюсків, ракоподібних і риб [5].
1.3 Антропогенний вплив на водні екосистеми
1.3.1 Органічне забруднення
1.3.1.1 Органічні речовини та їх кругообіг у водних екосистемах
Одним із найважливіших компонентів водного середовища, що визначає його екологічну якість, є наявність у воді органічних забруднень.
У процесі життєдіяльності гідробіонти виділяють у воду білки, амінокислоти, вуглеводи, сечовину, пурини, фосфати, амонійні сполуки тощо. Фактично у водному середовищі знаходяться всі ті органічні речовини, з яких побудовано тіло рослин і тварин. Крім того, органічні речовини надходять у водні об’єкти з атмосферними опадами, з поверхневим стоком, що формується на великих площах суходолу, з боліт, торф’яників, зрошувальних земель, промислових та комунально-побутових підприємств. Усі ці стоки привносять значну кількість різноманітних за своєю структурою і хімічним складом органічних речовин. За походженням органічні речовини поділяються на алохтонні, які надходять з площі водозбору, та автохтонні, що утворюються в самій водній екосистемі. Найбільшу масу органічної речовини створюють фітопланктон і макроліти в процесі фотосинтезу.
Значну кількість автохтонної органічної речовини становить детрит, або мертва органічна речовина, яка утворюється внаслідок розкладу залишків організмів рослинного і тваринного походження і містить також 4-5% бактерій.
Органічні речовини природних вод — це складні високомолекулярні сполуки типу білків, полісахаридів, ненасичених жирних кислот, амінів та інших, які знаходяться в розчиненому, завислому і диспергованому станах. Більшість з них є субстратом для масового розвитку бактеріального населення водних екосистем [8].
Найважливішим внутрішньоводоймним процесом утворення органічної речовини, з яким пов’язане і само забруднення водойм, є фотосинтез фітопланктону, фітобентосу та вищої водяної рослинності. Вважається, що в межах усього Світового океану внесок водоростей в утворення органічної речовини у 10 разів більший, ніж усіх інших груп організмів, не рахуючи бактерій. Останнім належить виключно важлива роль не тільки в розкладанні, а й в утворенні органічної речовини. У процесі функціонування водних екосистем значна частина розчиненої і завислої органічної речовини алохтонного і автохтонного походження виноситься із стоком води та розкладається в процесі деструкції (мінералізації).
Продукція і деструкція органічної речовини характеризують функціональний стан водних екосистем. Рівень і спрямованість продукційно-деструкційних процесів залежать перш за все від ступеню розвитку фітопланктону та умов його вегетації. У морських і континентальних водоймах утворення первинної органічної речовини пов’язане з життєдіяльністю планктонних і донних водоростей, макрофітів і епіфітів, які формують автотрофну ланку водних екосистем.
Синтезована органічна речовина є основою трофічної піраміди, по якій розподіляються потоки енергії у водних екосистемах. Саме органічна речовина автотрофних організмів забезпечує функціонування вищих (наступних) трофічних рівнів, біотичний кругообіг речовин і потік енергії в екосистемах. У кінцевому підсумку формується біологічна продуктивність водних екосистем. В залежності від рівня утворення первинної продукції їх поділяють на оліготрофні (малопродуктивні), мезотрофні (середньопродуктивні), евтотрофні (високопродуктивні) і гіперевтрофні (надмірно продуктивні) [5].
Для функціонування водних екосистем важливе значення має співвідношення первинної продукції органічної речовини (А — assimilation), при якому енергія накопичується, до деструкції (R — reduction) або сумарного дихання гідробіонтів (respiration), в процесі якого відбувається деструкція органічної речовини і розсіяння енергії. Підвищення біопродукційного потенціалу водних екосистем відбувається при евтрофікації.
Похожие работы
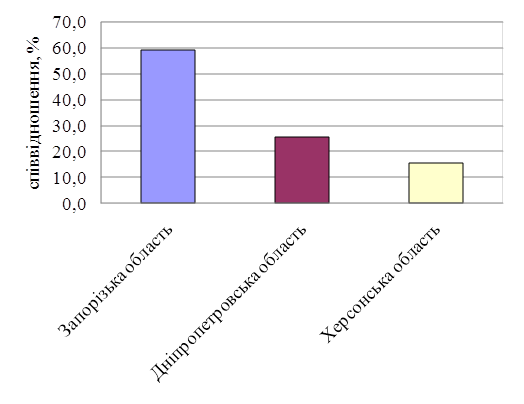
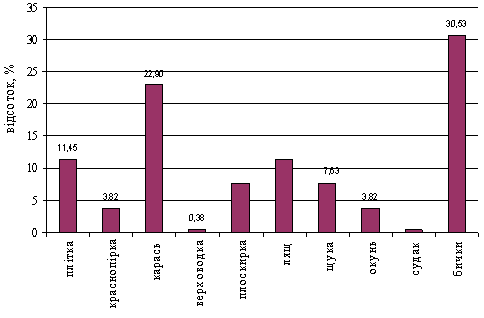
... види, як верховодка – 0,38%, краснопірка – 3,82% та інші (рисунок 3.4). Рисунок 3.4 – Видовий склад уловів риболовів-любителів на Каховському водосховищі у 2006-2009 роках 3.3 Ефективність засобів та знарядь любительського лову риби Оцінка впливу насадки, що використовується на інтенсивність лову поплавковою вудкою здійснювалась за результатами опитування 260 рибалок ...
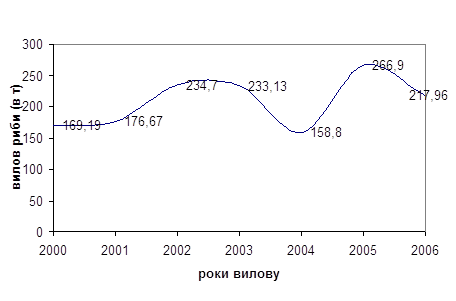

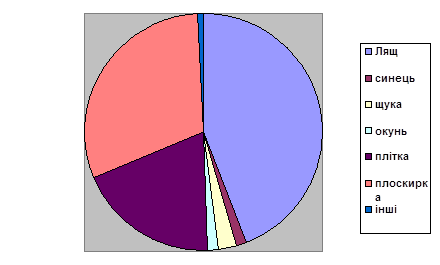
... міру намерзання льоду риба концентрується в ямах, з яких її вичерпують сачками. РОЗДІЛ 4. АНАЛІЗ ДИНАМІКИ ПОКАЗНИКІВ ВИЛОВУ РИБИ В ПРИРОДНИХ ВОДОЙМАХ 4.1 Природоохоронні заходи, які сприяють збільшенню цінних видів риби у річках Полісся Водойми мають визначену рибопродуктивність, що залежить від сукупності умов, зокрема від кормових ресурсів. У рибництві під природною рибопродуктивністю ...


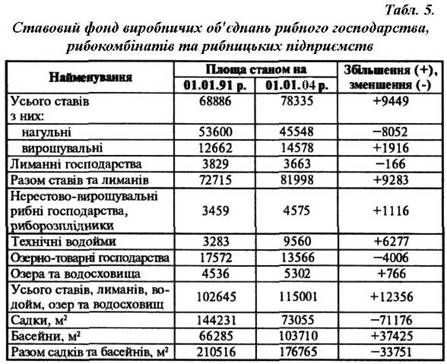
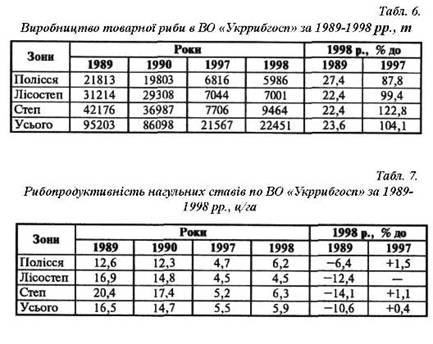
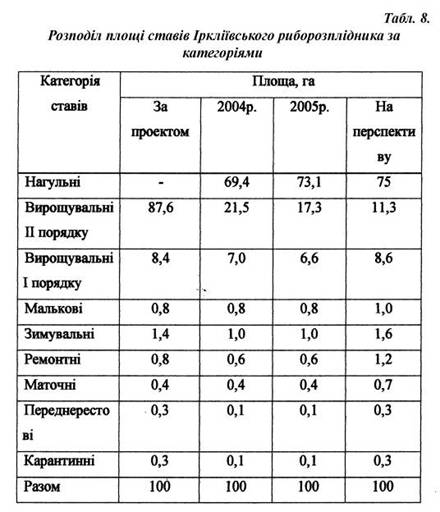
... матеріал та методика роботи. Дослідження проводили на базі Іркліївського риборозплідника рослиноїдних риб вздовж виробничої практики. З метою вивчення шляхів інтенсифікації отримання рибницької продукції в умовах Іркліївського риборозплідника рослиноїдних риб проводилось ознайомлення і вивчення наявних заходів інтенсифікації ведення рибогосподарства. Також проходило ознайомлення з запланованими ...
... ів зумовила зростаючу зацікавленість колгоспів, радгоспів, спеціалізованих рибогосподарських й наукових організацій до водойм різного походження, призначення та відомчої підпорядкованості в плані виробництва товарної риби. До цієї групи водойм належать малі водосховища, створені в результаті цілеспрямованої діяльності, що пов’язана з необхідністю перерозподілу строку або створення стабільних запас ...
0 комментариев