Навигация
2. Биокинетика
Один из основных разделов кинетики изучает кинетику биологических реакций; этот раздел принято называть биокинетикой. Биокинетика является пограничной наукой, возникшей на стыке биохимии и химической кинетики (см рис. 1). Выделение биокинетики в отдельную дисциплину неслучайно, оно логически оправдано и связано с исключительной значимостью кинетических процессов для всех живых организмов. Биокинетика — относительно молодая наука. Термин «Биокинетика» был введен И.В. Березиным и С.Д. Варфоломеевым в 1979 г. Традиционно в курсе биокинетики рассматриваются ферментативные реакции, процессы взаимодействия лигандов с рецепторами и процессы клеточного роста.
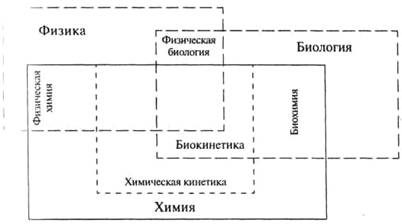
Рисунок 1. « Соотношение между основными естественнонаучными дисциплинами».
3. Движение клеток
При изучении живых систем часто говорится о движении: движутся хромосомы к полюсам клетки во время митоза, перемещаются вакуоли клеточных органелл, движется клеточная поверхность. Кроме того, в клетках растений и животных наблюдаются токи цитоплазмы (например, в растительных клетках или у амебы). Более того, отдельные клетки (свободноживущие одноклеточные организмы или специфические типы клеток в многоклеточных животных организмах) обладают способностью активно перемещаться, «ползать» (см. рис. 3.). Некоторые клетки имеют специализированные структуры, реснички или жгутики, которые позволяют им или самым перемещаться, или перемещать окружающую их жидкость. Наконец, у многоклеточных животных организмов есть специализированные клетки, мышечная работа которых позволяет производить различные движения органов, отдельных его частей и всего организма. В основе всех этих многочисленных двигательных реакций лежат общие молекулярные механизмы. Кроме того, наличие каких-либо двигательных аппаратов должно сочетаться и структурно связываться с существованием опорных, каркасных или скелетных внутриклеточных образований. Поэтому можно говорить (описывать и изучать) об опорно-двигательной системе клеток.
Само понятие о цитоскелете или скелетных компонентах цитоплазмы разных клеток было высказано Н.К.Кольцовым, выдающимся русским цитологом еще в начале XX века. К сожалению, они были забыты и только в конце 1950-х годов с помощью электронного микроскопа эта скелетная система была переоткрыта.
Огромный вклад в изучение цитоскелета внес метод иммунофлуоресценции, который помог разобраться в химии и динамике этого чрезвычайно важного компонента клетки. Цитоскелетные компоненты представлены нитевидными, неветвящимися белковыми комплексами, или филаментами (тонкими нитями).
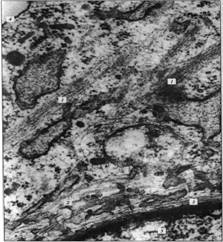
Рисунок 2. «Микрофотография элементов цитоскелета, полученная с помощью электронного микроскопа». 1 - пучки микрофиламентов; 2 - микротрубочки; 3 - промежуточные филаменты 4 плазматическая мембрана; 5 – ядро.

Рисунок 3. «Поляризованные движущиеся фибробласты в культуре ткани». 1 – ламеллоплазма; 2 — ядро
Существуют три системы филаментов, различающихся по химическому составу, ультраструктуре и функциональным свойствам. Самые тонкие нити — это микрофиламенты; их диаметр составляет около 6 мм, и состоят они в основном из белка актина. К другой группе нитчатых структур относятся микротрубочки, которые имеют диаметр 25 нм и состоят в основном из белка тубулина. Третья группа представлена промежуточными филаментами с диметром около 10 нм (промежуточным по сравнению с 6 и 25 нм), образующимися из разных, но родственных белков (рис. 2 и 4).
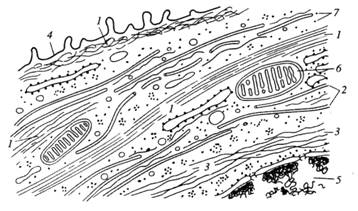
Рисунок 4. «Схематическое изображение цитоскелетных компонентов клеток». 1 – микрофиламенты; 2 – микротрубочки; 3 – промежуточные филаменты; 4 – плазмалитическая мембрана; 5 – ядро; 6 – митохондрии; 7 – рибосомы.
4. Системы подвижности эукариотических клеток
4.1 Микротрубочки
Микротрубочки предстали как особые внутриклеточные структуры (особенно внутри жгутика) благодаря применению методов электронной микроскопии. В 1946 г. Джакус и Холл продемонстрировали наличие одинаковых по диаметру трубчатых структур в ресничках Paramecium. Впоследствии трубчатые структуры обнаружили почти во всех клетках. Стали считать, что они характерны для эукариотичсского уровня организации. Однако микротубулярные структуры были обнаружены и у прокариот — в цитоплазме спирохет. Морфологически и биохимически эти микротрубочки подобны тем, которые наблюдаются в эукариотических клетках. Это открытие поддержало «экзогенную гипотезу» происхождения микротрубочек эукариот, согласно которой реснички и жгутики клеток высших организмов считаются приобретенными извне—путем симбиоза клеток, ранее не имевших жгутиков или ресничек, со спирохетами, содержавшими микротрубочки.
Микротрубочки представляют собой длинные полые цилиндры, наружный диаметр которых около 24 нм, а внутренний—15 нм. В большинстве клеток длина трубочек обычно не превышает нескольких микрон, хотя в некоторых специализированных клетках, например в моторных нейронах центральной нервной системы, трубочки могут быть длиной в несколько сантиметров. Стенка микротрубочки (около 5 нм толщиной) построена из продольно ориентированных протофибрилл. Протофибриллы состоят из глобулярных субъединиц (их очень хорошо видно в электронный микроскоп при негативном окрашивании), содержащих только один белок – тубулин (рис. 5). Помимо того, что микротрубочки имеют прямое отношение к подвижности клетки, они участвуют и в других процессах, более или менее связанных с подвижностью, например в поддержании формы клетки, во внутриклеточном транспорте веществ, в секреции клеточных продуктов, в движении хромосом при делении клетки и, возможно, в осуществлении сенсорных связей, а также в перемещении компонентов клеточной мембраны. Микротрубочки могут быть рассеяны по всей цитоплазме, а могут быть собраны в организованные структуры.

Рисунок 5. «Электронные, микрофотографии микротрубочек в краевых областях эритроцитов тритона»: а — продольный срез, на котором: видна линейная упаковка субъединиц в микротрубочках; б—поперечный срез; видно, что стенка каждой микротрубочки образована тринадцатью субъединицами. Негативное окрашивание. ×200 000.
Чтобы понять, как клетка движется с помощью микротрубочек, мы рассмотрим их организацию в ресничках жгутиках эукариотических клеток, в сократимом аксостиле некоторых жгутиконосцев, а также в микротубулярной сети цитоплазмы животных клеток.
4.1.1 Микротрубочки в ресничках и жгутикахЭукариотические реснички и жгутики представляют собой специализированные структуры, выступающие за пределы клеточной поверхности и способные двигаться. Эти два типа органелл идентичны по своей структуре; в функциональном отношении между ними обнаруживаются лишь незначительные, различия; Жгутики и реснички удается легко и в довольно больших количествах отделить от клеток, иногда просто энергичным встряхиванием клеточной суспензии. Это облегчило исследование их структуры и химического состава.
Структурная организация ресничек и жгутиков практически во всех исследовавшихся клетках отличается удивительным единообразием: центральную пару микротрубочек окружает кольцо из микротрубочек (обычно парных). Микротрубочки связаны между собой поперечными мостиками; вся конструкция называется аксонемой жгутика. Расположение микротрубочек и мостиков в аксонеме изучено в настоящее время довольно подробно; оно схематически представлено на рис. 6. В центре находится пара микротрубочек, окруженных общей оболочкой из белковых колец, плоскость которых наклонена к оси аксонемы. Стенка каждой из центральных микротрубочек образована тринадцатью продольными рядами (протофибриллами) белковых субъединиц. Эту центральную структуру, окружают девять расположенных по кольцу пар микротрубочек. В каждом дублете одна микротрубочка (основная или А) имеет полностью замкнутую стенку, состоящую из 13 протофибрилл (рис. 7); другая микротрубочка (микротрубочка В) имеет серповидное сечение и примыкает к микротрубочке А, так что ее стенка состоит из меньшего числа протофибрилл, обычно из 10. Каждая микротрубочка А соединена с микротрубочкой В соседнего дублета с помощью мостика из белка, называемого нексином. От обращенной внутрь поверхности микротрубочки А отходит по направлению к центру белковый выступ — так называемая радиальная спица. Вдобавок каждая микротрубочка А имеет двойной ряд боковых ручек (их длина около 14 нм), построенных из белка (динеина) и направленных к соседнему дублету. Все эти выступы расположены с определенной периодичностью относительно оси микротрубочки А и, как будет видно из дальнейшего, играют очень важную роль в генерации движения. Описанное выше строение аксонемы соответствует всей средней части жгутика, т. е. за исключением его концевой части и основания, где аксонема устроена несколько иначе. На самом конце центральная пара микротрубочек обычно длиннее, чем окружающие ее периферические дублеты, так что жгутик на конце заострен. Нередко микротрубочки В бывают короче микротрубочек А, и периферические дублеты вблизи конца жгутика переходят в одиночные трубочки (рис. 6). На одиночных микротрубочках А в этой области жгутика выступов нет.

Рисунок 6. «Схематическое изображение аксонемы жгутика». Показана взаимосвязь между элементами структуры на разных уровнях: а — на конце жгутика, б — в средней части и в — на уровне базального тела.

Рисунок 7. «Схематическое изображение расположения тубулиновых субъединиц в микротрубочке»: а — поперечный срез; б — вид сбоку.
Специализированная структура — базальное тело, — из которой исходит жгутик, находится в цитоплазме клетки у основания жгутика. Она может быть различна в клетках разных типов, однако при этом основные, общие черты сохраняются. В базальном теле ресничек Tetrahymena piriformis наряду с микротрубочками А и В имеются еще микротрубочки С (они образованы десятью протофибриллами), так что по периферии реснички располагаются не дублеты, а триплеты микротрубочек, Эти триплеты связаны между собой боковыми выступами (поперечными мостиками) и прикреплены к центральному кольцу радиальными спицами (рис. 6). Базальное тело реснички или жгутика по своей структуре, похоже на центриоль животных клеток. И базальное тело, и центриоль действуют как организаторы ассоциации микротрубочек; в первом случае/образуется ресничка или жгутик, во втором — мнтотическое веретено.
Еще одна интересная часть жгутика — это так называемый воротничок, кольцевой валик, окружающий основание жгутика там, где он выходит из клетки. В электронный микроскоп па поперечных срезах воротничка в этой области видны волокнистые соединения между периферическими дублетами жгутика и внутренними слоями клеточной мембраны. Они жестко закрепляют микротрубочки в клеточной мембране и участвуют в движении жгутика.
Подвижность ресничек и жгутиков обусловлена свойствами именно аксонемы, но не других компонентов этих органелл (т. е. клеточной мембраны, цитоплазмы и др.).
Похожие работы



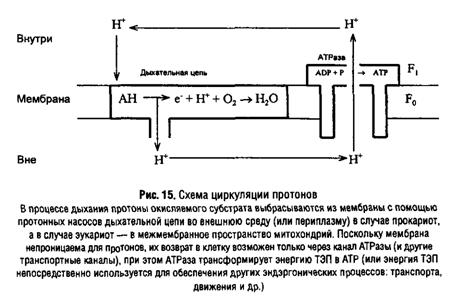
... подавления активности ферментов служит одним из способов расшифровки механизма их действия. Подходом к решению последней задачи является изучение специфичности действия ферментов. В свою очередь, это требует корректного измерения кинетических параметров в присутствии изучаемого аналога субстрата. Рассмотрим способы определения характера взаимоотношений субстратов, их аналогов и ингибиторов ...

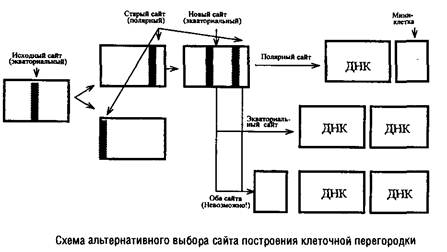
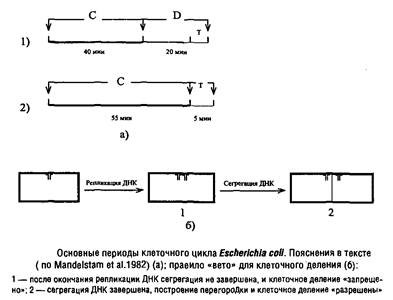
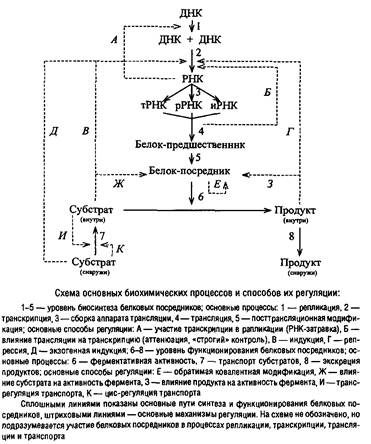
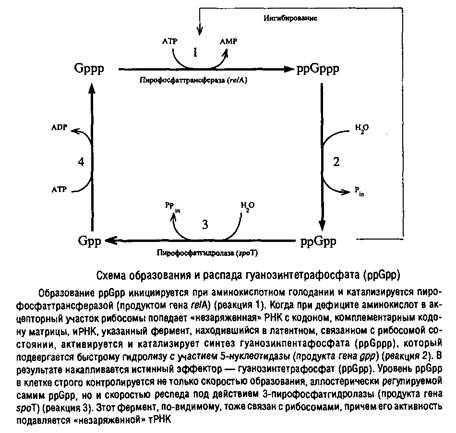
... Escherichia coti около 1 мкм за 25 с. Кроме того, способ сегрегации может определяться скоростью роста микроорганизма: у медленно растущих клеток Escherichia coii он близок к биполярному, а у быстро растущих становится днсперсивным. Построение клеточной перегородки В изучении механизмов регуляции данного этапа клеточного цикла важную роль сыграли специфические мутанты, особенно мутанты ...
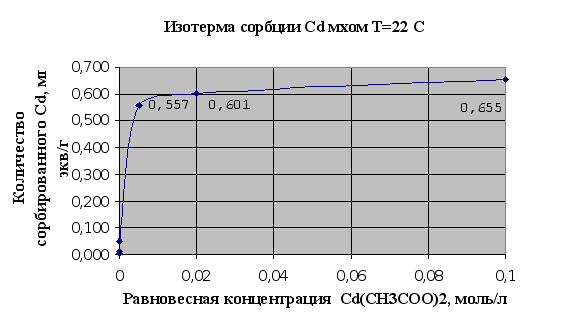
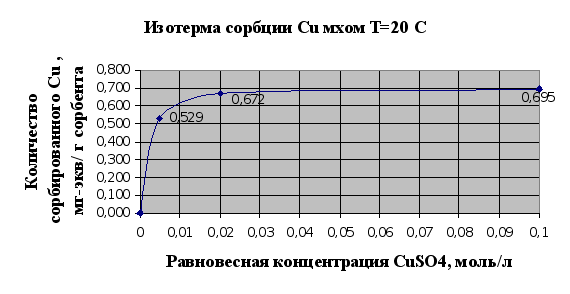
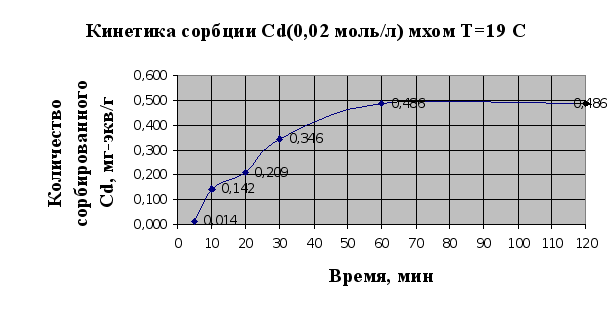

... Листов Пров. 1 2 Консульт. БГТУ 7140607 2004 Н. контр. Утв. Целью данной дипломной работы является изучение сорбционных свойств мха по отношению к микроорганизмам и тяжелым металлам, а также изучение сорбционной способности системы «мох-микроорганизмы» и выявление доли участия каждого компонента этой ...
... переломов ослабленных костей / Тр. конгресса Человек и его здоровье. СПб, 1999 - с. 55. 44. Воложин А.И., Курдюмов С.Г., Орловский В.П., Баринов С.М. и др. Создание нового поколения биосовместимых материалов на основе фосфатов кальция для широкого применения в медицинской практике // Технологии живых систем. 2004. Т.1, №.1. С. 41-56. 45. Безруков В.М., Григорян А.С. Гидроксилапатит как ...
0 комментариев