Навигация
Приспособление у растений
38470
знаков
0
таблиц
0
изображений
2. Приспособление у растений
Холодостойкость растенийУстойчивость растений к низким температурам подразделяют на холодостойкость и морозоустойчивость. Под холодостойкостью понимают способность растений переносить положительные температуры несколько выше Оє С. Холодостойкость свойственна растениям умеренной полосы (ячмень, овес, лен, вика и др.). Тропические и субтропические растения повреждаются и отмирают при температурах от 0є до 10є С (кофе, хлопчатник, огурец и др.). Для большинства же сельскохозяйственных растений низкие положительные температуры негубительны. Связано это с тем, что при охлаждении ферментативный аппарат растений не расстраивается, не снижается устойчивость к грибным заболеваниям и вообще не происходит заметных повреждений растений.
Степень холодостойкости разных растений неодинакова. Многие растения южных широт повреждаются холодом. При температуре 3 °С повреждаются огурец, хлопчатник, фасоль, кукуруза, баклажан. Устойчивость к холоду у сортов различна. Для характеристики холодостойкости растений используют понятие температурный минимум, при котором рост растений прекращается. Для большой группы сельскохозяйственных растений его величина составляет 4 °С. Однако многие растения имеют более высокое значение температурного минимума и соответственно они менее устойчивы к воздействию холода.
Приспособление растений к низким положительным температурам.Устойчивость к низким температурам — генетически детерминированный признак. Холодостойкость растений определяется способностью растений сохранять нормальную структуру цитоплазмы, изменять обмен веществ в период охлаждения и последующего повышения температуры на достаточно высоком уровне.
Морозоустойчивость растенийМорозоустойчивость — способность растений переносить температуру ниже О °С, низкие отрицательные температуры. Морозоустойчивые растения способны предотвращать или уменьшать действие низких отрицательных температур. Морозы в зимний период с температурой ниже -20 °С обычны для значительной части территории России. Воздействию морозов подвергаются однолетние, двулетние и многолетние растения. Растения переносят условия зимы в различные периоды онтогенеза. У однолетних культур зимуют семена (яровые растения), раскустившиеся растения (озимые), у двулетних и многолетних — клубни, корнеплоды, луковицы, корневища, взрослые растения. Способность озимых, многолетних травянистых и древесных плодовых культур перезимовывать обусловливается их достаточно высокой морозоустойчивостью. Ткани этих растений могут замерзать, однако растения не погибают.
Замерзание растительных клеток и тканей и происходящие при этом процессы.Способность растений переносить отрицательные температуры определяется наследственной основой данного вида растений, однако морозоустойчивость одного и того же растения зависит от условий, предшествующих наступлению морозов, влияющих на характер льдообразования. Лед может образовываться как в протопласте клетки, так и в межклеточном пространстве. Не всякое образование льда приводит клетки растения к гибели.
Постепенное снижение температуры со скоростью 0,5—1 °С/ч приводит к образованию кристаллов льда прежде всего в межклеточниках и первоначально не вызывают гибели клеток. Однако последствия этого процесса могут быть губительными для клетки. Образование льда в протопласте клетки, как правило, происходит при быстром понижении температуры. Происходит коагуляция белков протоплазмы, кристаллами образовавшегося в цитозоле льда повреждаются клеточные структуры, клетки погибают. Убитые морозом растения после оттаивания теряют тургор, из их мясистых тканей вытекает вода.
Морозоустойчивые растения обладают приспособлениями, уменьшающими обезвоживание клеток. При понижении температуры у таких растений отмечаются повышение содержания Сахаров и других веществ, защищающих ткани (криопротекторы), это прежде всего гидрофильные белки, моно- и олигосахариды; снижение оводненности клеток; увеличение количества полярных липидов и снижение насыщенности их жирнокислотных остатков; увеличение количества защитных белков.
На степень морозоустойчивости растений большое влияние оказывают сахара, регуляторы роста и другие вещества, образующиеся в клетках. В зимующих растениях в цитоплазме накапливаются сахара, а содержание крахмала снижается. Влияние сахаров на повышение морозоустойчивости растений многосторонне. Накопление Сахаров предохраняет от замерзания большой объем внутриклеточной воды, заметно уменьшает количество образующегося льда.
Свойство морозоустойчивости формируется в процессе онтогенеза растения под влиянием определенных условий среды в соответствии с генотипом растения, связано с резким снижением темпов роста, переходом растения в состояние покоя.
Жизненный цикл развития озимых, двулетних и многолетних растений контролируется сезонным ритмом светового и температурного периодов. В отличие от яровых однолетних растений они начинают готовиться к перенесению неблагоприятных зимних условий с момента остановки роста и затем в течение осени во время наступления пониженных температур.
Зимостойкость растений Зимостойкость как устойчивость к комплексу неблагоприятных факторов перезимовки.Непосредственное действие мороза на клетки - не единственная опасность, угрожающая многолетним травянистым и древесным культурам, озимым растениям в течение зимы. Помимо прямого действия мороза растения подвергаются еще ряду неблагоприятных факторов. В течение зимы температура может существенно колебаться. Морозы нередко сменяются кратковременными и длительными оттепелями. В зимнее время нередки снежные бури, а в бесснежные зимы в более южных районах страны - и суховеи. Все это истощает растения, которые после перезимовки выходят сильно ослабленными и в последующем могут погибнуть.
Особенно многочисленные неблагоприятные воздействия испытывают травянистые многолетние и однолетние растения. На территории России в неблагоприятные годы гибель посевов озимых зерновых достигает 30—60 %. Погибают не только озимые хлеба, но и многолетние травы, плодовые и ягодные многолетние насаждения. Кроме низких температур озимые растения повреждается и гибнут от ряда других неблагоприятных факторов в зимнее время и ранней весной: выпревания, вымокания, ледяной корки, выпирания, повреждения от зимней засухи.
Выпревание, вымокание, гибель под ледяной коркой, выпирание, повреждение от зимней засухи.Выпревание. Среди перечисленных невзгод первое место занимает выпревание растений. Гибель растений от выпревания наблюдается преимущественно в теплые зимы с большим снеговым покровом, который лежит 2—3 месяца, особенно если снег выпадает на мокрую и талую землю. Исследования показали, что причина гибели озимых от выпревания — истощение растений. Находясь под снегом при температуре около О °С в сильно увлажненной среде, почти полной темноте, т. е. в условиях, при которых процесс дыхания идет достаточно интенсивно, а фотосинтез исключен, растения постепенно расходуют сахара и другие запасы питательных веществ, накопленные в период прохождения первой фазы закаливания, и погибают от истощения (содержание Сахаров в тканях уменьшается с 20 до 2—4 %) и весенних заморозков. Такие растения весной легко повреждаются снежной плесенью, что также приводит к их гибели.
Вымокание. Вымокание проявляется преимущественно весной в пониженных местах в период таяния снега, реже во время длительных оттепелей, когда на поверхности почвы накапливается талая вода, которая не впитывается в замершую почву и может затопить растения. В этом случае причиной гибели растений служит резкий недостаток кислорода (анаэробные условия - гипоксия). У растений, оказавшихся под слоем воды, нормальное дыхание прекращается из-за недостатка кислорода в воде и почве. Отсутствие кислорода усиливает анаэробное дыхание растений, в результате чего могут образоваться токсичные вещества и растения погибают от истощения и прямого отравления организма.
Гибель под ледяной коркой. Ледяная корка образуется на полях в районах, где частые оттепели сменяются сильными морозами. Действие вымокания в этом случае может усугубляться. При этом происходит образование висячих или притертых (контактных) ледяных корок. Менее опасны висячие корки, так как они образуются сверху почвы и практически не соприкасаются с растениями; их легко разрушить катком.
При образовании же сплошной ледяной контактной корки растения полностью вмерзают в лед, что ведет к их гибели, так как и без того ослабленные от вымокания растения подвергаются очень сильному механическому давлению.
Выпирание. Повреждение и гибель растений от выпирания определяются разрывами корневой системы. Выпирание растений наблюдается, если осенью морозы наступают при отсутствии снежного покрова или если в поверхностном слое почвы мало воды (при осенней засухе), а также при оттепелях, если снеговая вода успеет всосаться в почву. В этих случаях замерзание воды начинается не с поверхности почвы, а на некоторой глубине (где есть влага). Образующаяся на глубине прослойка льда постепенно утолщается за счет продолжающегося поступления воды по почвенным капиллярам и поднимает (выпирает) верхние слои почвы вместе с растениями, что приводит к обрыву корней растений, проникших на значительную глубину.Повреждения от зимней засухи. Устойчивый снеговой покров предохраняет озимые злаки от зимнего высыхания. Однако они в условиях бесснежной или малоснежной зимы, как и плодовые деревья и кустарники, в ряде районов России часто подвергаются опасности чрезмерного иссушения постоянными и сильными ветрами, особенно в конце зимы при значительном нагреве солнцем. Дело в том, что водный баланс растений складывается зимой крайне неблагоприятно, так как поступление воды из замерзшей почвы практически прекращается.
Для уменьшения испарения воды, неблагоприятного действия зимней засухи плодовые древесные породы образуют на ветвях мощный слой пробки, сбрасывают на зиму листья.
ЯровизацияФотопериодические реакции на сезонные изменения длины дня имеют значение для периодичности цветения многих видов как умеренных, так и тропических областей. Однако следует отметить, что среди видов умеренных широт, проявляющих фотопериодические реакции, относительно мало весеннецветущих, хотя мы постоянно сталкиваемся с тем, что значительное число «цветов цветет весной», и многие из таких весеннецветущих форм, например Ficaria verna, первоцвет (Primula vutgaris), фиалки (виды рода Viola) и т. д., проявляют выраженное сезонное поведение, оставаясь вегетативными оставшуюся часть года после обильного весеннего цветения. Можно предположить, что весеннее цветение -реакция на короткие дни зимой, но для многих видов, это, по-видимому, не так.
Конечно, длина дня не является единственным внешним фактором, изменяющимся в течение года. Ясно, что и температура также характеризуется четко выраженными сезонными изменениями, особенно в умеренных областях, хотя в отношении этого фактора наблюдаются значительные колебания, как ежедневные, так и ежегодные. Мы знаем, что сезонные изменения температуры, так же как и изменения длины дня, оказывают существенное влияние на цветение многих видов растений.
Типы растений, требующих охлаждения для перехода к цветению.Было установлено, что многие виды, в том числе озимые однолетние, а также двулетние и многолетние травянистые растения, нуждаются в охлаждении для перехода к цветению.
Известно, что озимые однолетники и двулетники представляют собой монокарпические растения, которые требуют яровизации,- они остаются вегетативными во время первого вегетативного сезона и цветут следующей весной или ранним летом в ответ на период охлаждения, получаемый зимой. Необходимость охлаждения двулетних растений для индукции цветения была экспериментально продемонстрирована на ряде видов, таких, как свекла (Beta vulgaris), сельдерей (Apiutn graveolens), капуста и другие культивируемые сорта рода Brassiса, колокольчик (Campanula medium), лунник (Lunaria biennis), наперстянка (Digitalis purpurea) и другие. Если растения наперстянки, которые в нормальных условиях ведут себя как двулетники, т. е. зацветают на второй год после прорастания, содержать в оранжерее, они могут оставаться вегетативными несколько лет. В районах с мягкой зимой капуста в течение нескольких лет может расти в открытом грунте без «образования стрелки» (т. е. цветения) весной, что обычно происходит в районах с холодной зимой. Такие виды обязательно требуют яровизации, однако у ряда других видов цветение ускоряется при воздействии на них холодом, но может наступать и без яровизации; к таким видам, проявляющим факультативную потребность в холоде, относятся салат (Lactuca saiiva), шпинат (Spinacia oleracea) и позднецветущие сорта гороха (Pistim sa-tivum).
Так же как и двулетние, многие многолетние виды нуждаются в воздействии холодом и не зацветают без ежегодного зимнего охлаждения. Из обычных многолетних растений в холодовом воздействии нуждаются первоцвет (Primula vulgaris), фиалки (Viola spp.), лакфиоль (Cheiranthus cheirii и С. allionii), левкой (Mathiola incarna), некоторые сорта хризантем (Chrisant-hemum morifolium), виды рода Aster, турецкая гвоздика (Dianthus), плевел (Lolium perenne). Многолетние виды требуют переяровизации каждую зиму.
Вполне вероятно, что и у других весеннецветущих многолетников можно обнаружить потребность в охлаждении. Весенне-цветущие луковичные растения, такие, как нарциссы, гиацинты, пролески (Endymion nonscriptus), крокусы и т. д. не требуют охлаждения для заложения цветков, поскольку примордий цветка заложился в луковице предыдущим летом, но их рост в значительной степени зависит от температурных условий. Например, у тюльпана началу цветения благоприятствуют относительно высокие температуры (20°С), но для удлинения стебля и роста листьев оптимальной температурой вначале является 8-9 °С с последовательным повышением на более поздних стадиях до 13, 17 и 23°С. Аналогичные реакции на температуру характерны для гиацинтов и нарциссов.
У многих видов заложение цветка происходит не во время самого периода охлаждения и начинается лишь после того, как растение подверглось действию более высоких температур, следующих за охлаждением.
Таким образом, хотя при низких температурах метаболизм у большинства растений значительно замедляется, не вызывает сомнения, что яровизация включает активные физиологические процессы, природа которых пока еще совершенно неизвестна.
Жароустойчивость растенийЖароустойчивость (жаровыносливость) - способность растений переносить действие высоких температур, перегрев. Это генетически обусловленный признак. Виды растений различаются по выносливости к высоким температурам.
По жароустойчивости выделяют три группы растений.
Жаростойкие - термофильные сине-зеленые водоросли и бактерии горячих минеральных источников, способные переносить повышение температуры до 75-100 °С. Жароустойчивость термофильных микроорганизмов определяется высоким уровнем метаболизма, повышенным содержанием РНК в клетках, устойчивостью белка цитоплазмы к тепловой коагуляции.
Жаровыносливые - растения пустынь и сухих мест обитания (суккуленты, некоторые кактусы, представители семейства Толстянковые), выдерживающие нагревание солнечными лучами до 50-65єС. Жароустойчивость суккулентов во многом определяется повышенными вязкостью цитоплазмы и содержанием связанной воды в клетках, пониженным обменом веществ.
Нежаростойкие - мезофитные и водные растения. Мезофиты открытых мест переносят кратковременное действие температур 40-47 єС, затененных мест - около 40-42 °С, водные растения выдерживают повышение температуры до 38-42 °С. Из сельскохозяйственных наиболее жаровыносливы теплолюбивые растения южных широт (сорго, рис, хлопчатник, клещевина и др.).
Многие мезофиты переносят высокую температуру воздуха и избегают перегрева благодаря интенсивной транспирации, снижающей температуру листьев. Более жаростойкие мезофиты отличаются повышенной вязкостью цитоплазмы и усиленным синтезом жаростойких белков-ферментов.
Растения выработали систему морфологических и физиологических приспособлений, защищающих их от тепловых повреждений: светлую окраску поверхности, отражающую инсоляцию; складывание и скручивание листьев; опушения или чешуйки, защищающие от перегрева глубжележащие ткани; тонкие слои пробковой ткани, предохраняющие флоэму и камбий; большую толщину кутикулярного слоя; высокое содержание углеводов и малое — воды в цитоплазме и др.
На тепловой стресс растения очень быстро реагируют индуктивной адаптацией. К воздействию высоких температур они могут подготовиться за несколько часов. Так, в жаркие дни устойчивость растений к высоким температурам после полудня выше, чем утром. Обычно эта устойчивость временная, она не закрепляется и довольно быстро исчезает, если становится прохладно. Обратимость теплового воздействия может составлять от нескольких часов до 20 дней. В период образования генеративных органов жаростойкость однолетних и двулетних растений снижается.
Засухоустойчивость растенийОбычным явлением для многих регионов России и государств СНГ стали засухи. Засуха - это длительный бездождливый период, сопровождаемый снижением относительной влажности воздуха, влажности почвы и повышением температуры, когда не обеспечиваются нормальные потребности растений в воде. На территории России имеются регионы неустойчивого увлажнения с годовым количеством осадков 250-500 мм и засушливые, с количеством осадков менее 250 мм в год при испаряемости более 1000 мм.
Засухоустойчивость - способность растений переносить длительные засушливые периоды, значительный водный дефицит, обезвоживание клеток, тканей и органов. При этом ущерб урожая зависит от продолжительности засухи и ее напряженности. Различают засуху почвенную и атмосферную.
Почвенная засуха вызывается длительным отсутствием дождей в сочетании с высокой температурой воздуха и солнечной инсоляцией, повышенным испарением с поверхности почвы и транспирацией, сильными ветрами. Все это приводит к иссушению корнеобитаемого слоя почвы, снижению запаса доступной для растений воды при пониженной влажности воздуха. Атмосферная засуха характеризуется высокой температурой и низкой относительной влажностью воздуха (10-20 %). Жесткая атмосферная засуха вызывается перемещением масс сухого и горячего воздуха - суховея. К тяжелым последствиям приводит мгла, когда суховей сопровождается появлением в воздухе почвенных частиц (пыльные бури).
Атмосферная засуха, резко усиливая испарение воды с поверхности почвы и транспирацию, способствует нарушению согласованности скоростей поступления из почвы в надземные органы воды и потери ее растением, в результате растение завядает. Однако при хорошем развитии корневой системы атмосферная засуха не причиняет растениям большого вреда, если температура не превышает переносимый растениями предел. Продолжительная атмосферная засуха в отсутствие дождей приводит к почвенной засухе, которая более опасна для растений.
Засухоустойчивость обусловлена генетически определенной приспособленностью растений к условиям места обитания, а также адаптацией к недостатку воды. Засухоустойчивость выражается в способности растений переносить значительное обезвоживание за счет развития высокого водного потенциала тканей при функциональной сохранности клеточных структур, а также за счет адаптивных морфологических особенностей стебля, листьев, генеративных органов, повышающих их выносливость, толерантность к действию длительной засухи.
Типы растений по отношению к водному режимуРастения засушливых областей называются ксерофитами (от греческого хеrоs - сухой). Они способны в процессе индивидуального развития приспосабливаться к атмосферной и почвенной засухе. Характерные признаки ксерофитов — незначительные размеры их испаряющей поверхности, а также небольшие размеры надземной части по сравнению с подземной. Ксерофиты - это обычно травы или низкорослые кустарники. Их делят на несколько типов. Приводим классификацию ксерофитов по П. А. Генкелю.
Суккуленты — очень стойкие к перегреву и устойчивые к обезвоживанию, во время засухи они не испытывают недостатка воды, потому что содержат большое количество ее и медленно расходуют. Корневая система у них разветвлена во все стороны в верхних слоях почвы, благодаря чему в дождливые периоды растения быстро всасывают воду. Это кактусы, алоэ, очиток, молодило.
Эвксерофиты — жаростойкие растения, которые хорошо переносят засуху. К этой группе относятся такие степные растения, как вероника сизая, астра мохнатая, полынь голубая, арбуз колоцинт, верблюжья колючка и др. У них незначительная транспирация, высокое осмотическое давление, цитоплазма отличается высокой эластичностью и вязкостью, корневая система очень разветвлена, и основная ее масса размещена в верхнем слое почвы (50-60 см). Эти ксерофиты способны сбрасывать листья и даже целые ветви.
Гемиксерофиты, или полуксерофиты - это растения, которые неспособны переносить обезвоживание и перегрев. Вязкость и эластичность протопласта у них незначительная, отличается высокой транспирацией, глубокой корневой системой, которая может достигать подпочвенной воды, что обеспечивает бесперебойное снабжение растения водой. К этой группе относятся шалфей, резак обычный и др.
Стипаксерофшпы - это ковыль, тырса и другие узколистные степные злаки. Они устойчивы к перегреву, хорошо используют влагу кратковременных дождей. Выдерживают лишь кратковременную нехватку воды в почве.
Пойкилоксерофиты - растения, не регулирующие своего водного режима. Это в основном лишайники, которые могут высыхать до воздушно-сухого состояния и снова проявлять жизнедеятельность после дождей.
Гигрофиты (от греческого hihros - влажный). У растений, относящихся к этой группе, нет приспособлений, ограничивающих расход воды. Для гигрофитов характерны сравнительно большие размеры клеток, тонкостенная оболочка, слабоодревесневшие стенки сосудов, древесных и лубяных волокон, тонкая кутикула и малоутолщенные внешние стенки эпидермиса, большие устьица и незначительное количество их на единицу поверхности, большая листовая пластинка, плохо развитые механические ткани, редкая сеть жилок в листе, большая кутикулярная транспирация, длинный стебель, недостаточно развитая корневая система. По строению гигрофиты приближаются к теневыносливым растениям, но имеют своеобразную гигроморфную структуру. Незначительный недостаток воды в почве вызывает быстрое завядание гигрофитов. Осмотическое давление клеточного сока в них невысокое. К ним относятся манник, багульник, брусника, лох.
По условиям произрастания и особенностям строения к гигрофитам очень близки растения с частично или полностью погруженными в воду или плавающими на ее поверхности листьями, которые называются гидрофитами.
Мезофиты (от греческого mesos - средний, промежуточный). Растения этой экологической группы произрастают в условиях достаточного увлажнения. Осмотическое давление клеточного сока у мезофитов 1-1,5 тыс. кПа. Они легко завядают. К мезофитам относятся большинство луговых злаков и бобовых -пырей ползучий, лисохвост луговой, тимофеевка луговая, люцерна синяя и др. Из полевых культур твердые и мягкие пшеницы, кукуруза, овес, горох, соя, сахарная свекла, конопля, почти все плодовые (за исключением миндаля, винограда), многие овощные культуры (морковь, помидорыи др.).
Транспирирующие органы – листья отличаются значительной пластичностью; в зависимости от условий произрастания в их строении наблюдаются довольно большие отличия. Даже листья одного растения при разном водоснабжении и освещении имеют различия в строении. Установлены определенные закономерности в строении листьев в зависимости от расположения их на растении.
В. Р. Заленский обнаружил изменения в анатомическом строении листьев по ярусам. Он установил, что у листьев верхнего яруса наблюдаются закономерные изменения в сторону усиления ксероморфизма, т. е. происходит образование структур, повышающих засухоустойчивость этих листьев. Листья, расположенные в верхней части стебля, всегда отличаются от нижних, а именно: чем выше расположен лист на стебле, тем меньше размеры его клеток, большее количество устьиц и меньше их размеры, большее количество волосков на единицу поверхности, гуще сеть проводящих пучков, сильнее развита палисадная ткань. Все эти признаки характеризуют ксерофилию, т. е. образование структур, способствующих повышению засухоустойчивости.
С определенной анатомической структурой связаны и физиологические особенности, а именно: верхние листья отличаются более высокой ассимиляционной способностью и более интенсивной транспирацией. Концентрация сока в верхних листьях также более высокая, в связи с чем может происходить оттягивание воды верхними листьями от нижних, засыхание и отмирание нижних листьев. Структура органов и тканей, повышающая засухоустойчивость растений, называется ксероморфизмом. Отличительные особенности в структуре листьев верхнего яруса объясняются тем, что они развиваются в условиях несколько затрудненного водоснабжения.
Для уравнения баланса между поступлением и расходом воды в растении образовалась сложная система анатомо-физиологических приспособлений. Такие приспособления наблюдаются у ксерофитов, гигрофитов, мезофитов.
Результаты исследований показали, что приспособительные свойства у засухоустойчивых форм растений возникают под влиянием условий их существования.
ЗАКЛЮЧЕНИЕУдивительная гармония живой природы, ее совершенство создаются самой природой: борьбой за выживание. Формы приспособлений у растений и животных бесконечно разнообразны. Весь животный и растительный мир со времени своего появления совершенствуется по пути целесообразных приспособлений к условиям обитания: к воде, к воздуху, солнечному свету, силы тяжести и т.д.
ЛИТЕРАТУРА
1. Володько И.К. ''Микроэлементы и устойчивость растений к неблагоприятным условиям'', Минск, Наука и техника, 1983г.
2. Горышина Т.К. ''Экология растений'', уч. Пособие для ВУЗов, Москва, В. школа, 1979г.
3. Прокофьев А.А. ''Проблемы засухоустойчивости растений'', Москва, Наука, 1978г.
4. Сергеева К.А. ''Физиологические и биохимические основы зимостойкости древесных растений'', Москва, Наука, 1971г
5. Культиасов И.М. Экология растений. - М.: Изд-во московского ун-та, 1982
Похожие работы
... лежат наследственно закрепленные эндогенные циклы физиологических процессов с периодом, близким к 24 часам. Это циркадианные (циркадные) ритмы. Цирканнуальные ритмы - годовые ритмы. Глава 2.КЛИМАТ Волгоградская область расположена на юго-востоке Русской равнины, вдали от океанов и морей. Поэтому климат области континентальный, с холодной, малоснежной зимой и продолжительным, жарким, сухим летом ...
... . У засухоустойчивых растений эти показатели будут выше. В селекционной работе используют такой показатель, как содержание статолитного крахмала в корневом чехлике. Генетически обусловленным признаком засухоустойчивости растений является способность их вегетативных органов (особенно листьев) накапливать во время засухи пролин. При этом концентрация пролина увеличивается в 10—100 раз. В пролине ...
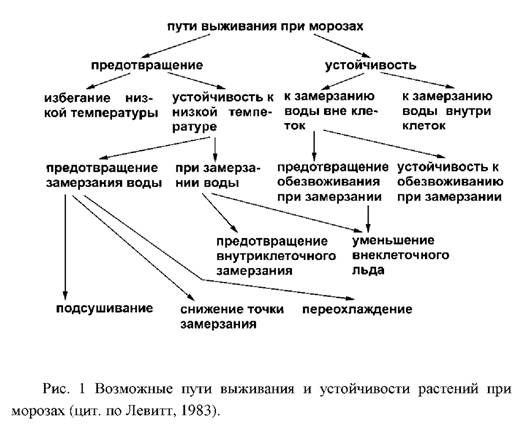
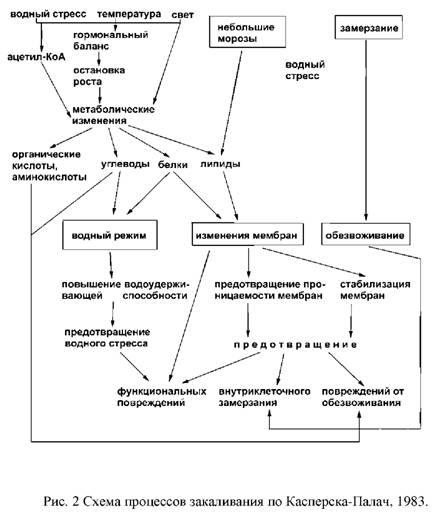
... и выбранные для контроля скорости роста при низко температуре и степени зимнего закаливания, были значительно менее устойчивы, чем норвежские популяции. Степень раззакаливания растени увеличивалась с повышением температуры. В условиях действия низко температуры северный экотип из Bodo был более устойчив к раззакаливанию, чем AberHerald. Тем не менее, при 18°C абсолютны уровень раззакаливания у ...



... фоне аллелопатических влияний сорных растений 4.3 Флористический состав, обилие и жизненное состояние сорных растений Исследования сорных растений в зерновом агрофитоценозе показали, что их сообщество включало до 89 видов, принадлежащих к 78 родам из 26 семейств. Сегетальная флора лесостепи Приобья при почвозащитной системе земледелия представлена преимущественно антропохорами (56.7%), на ...
0 комментариев