Африка. На африканском континенте эректус существовал менее 1 млн. лет назад (1.6/ 1.5- 0.3 млн. лет назад). Древнейшие бесспорные эректусы Восточной Африки обнаружены в Кении. Это череп 3733 из верхнего горизонта формации Кооби Фора древностью 1.5 млн. лет и находка почти полного скелета мальчика-подростка но стоянке нериокотоме-III( западный берег оз. Туркана), абсолютный возраст которой 1.6 млн. лет. Скелет принадлежал высокорослому архантропу с длиной тела (взрослого) до 180 см.; зубы очень сходны с зубами синантропа. К данным представителям таксона Н. Еrectus и иногда относят и череп 1470, а также некоторые кости посткраниального скелета (бедренная, тазовая) из Кооби Фора древностью около 12.89 млн. лет назад.
В ряде восточно- и, возможно, также североафриканских местонахождений прослеживается хронологическая, а подчас и морфологическая преемственность более ранних и поздних гоминид. Такая преемственность прослеживается между хабилисами и эректусами в Танзании (Олдувай), Кении (Кооби Фора) и Эфиопии (Омо). В Омо с помощью трех хронологических шкал (радиометрической, палеомагнитной и биостратиграфической) выявлено отчетливое “вертикальное” распределение гоминид рода Ноmо: Н. habilis с датировкой 2- 1.6 млн лет предшествует Н. Еrectus древностью около 1.2 млн. лет ; этот ряд замыкается архаическим сапиенсом, предполагаемый возраст которого примерно 0.1 млн. лет
Следовательно, около 100 000 лет назад в Омо (формация Кибиш) уже существовали гоминиды вполне сапиентног7о облика с округлым затылком, высоким сводом черепа, умерено выраженным рельефом лобной области, выступающим подбородком. Интересно, что в этом же местонахождении встречаются и более примитивные черепа (Омо II), обнаруживающие при современной емкости некоторые черты сходства с поздними африканскими эректусами или архаическими палеоанторопами , такими как Брохен Хилл (Замбия) или Хопфилд (ЮАР). Для всех этих гоминид характерна сильно изогнутая затылочная кость с развитым поперечным валиком . Если бы находки Омо I и Омо были сделаны в разных местах , то вполне возможно, что их сочли бы представителями различных филетических линий гоминид. И это далеко не единственный пример морфологического разнообразия в одном и том же локусе: изменчивость популяций ископаемого человека нередко была очень высокой. Существует точка зрения, что между всеми названными выше гоминидами Восточной и Южной Африки нет жесткой границы, их можно связать переходными формами как с более ранними находками, так и с “настоящим” протоавстралопидным сапиенсом из Флорисбада (ЮАР), имеющим древность около 40 000 лет.
Можно назвать еще несколько примеров раннего проявления сапиентации в Африке . Среди них ряд верхнеплейстоценовых гоминид из Восточной Африки, например, череп из Летоли (отложения Нгалоби) в Танзании древностью примерно 120 000 лет, сочетающий современные и архаические черты и обнаруживающий сходство с ранним африканским сапиенсом из Омо. К числу архаических сапиенсов периода 0.2- 0.1 млн. лет, по-видимому, относится и недавняя (1986год) находка (правда, не в первичном залегании) очень минерализованного черепа с емкостью 1300-1400см. куб. из области Западная Туркана в Кении (Элие-Спринге). Прослеживается сходство с группой Брокен-Хилл-Хопфилд. Особый интерес представляют некоторые материалы из района Восточная Туркана, прежде всего “челюстью 730” глубокой древности, но с такой поздней сапиентной специализацией, как подбородочный выступ. В Южной Африке известны скелетные остатки сапиенса а хорошо выраженным подбородком , датируемые 115 000-80 000 лет назад.
Возможна хронологическая и морфологическая преемственность гоминид и из североафриканского региона. Это атлантропы и близкие к ним формы из Алжира и Марокко (Тернифина, Сидиабдеррахман, Томас, Рабат, Сале, Темара) древностью 360 000-115 000 лет назад. Характерны некоторые архаические особенности: небольшая емкость черепов, массивный жевательный аппарат. Эволюционная преемственность прослеживается от этой группы к ископаемым африканским неоантропам (Афалу, Тафоральт и др.), а возможно, и к некоторым ранним европейским гоминидам (Мауэр, Монтморен). Высказывается также гипотеза о генеалогической связи африканских архантропов с местными палеонатропами, как например, Джебел Ирхуд их Морокко, которые послужили предками архаических кроманьонцев. Последние в период вюрмского оледенения переселились в Европу, где и дали начало кроманьонской расе (Ферембах, 1985).
Все эти данные подтверждают мнение, что процессы сапиентации в Африке действительно имеют весьма давние и обширные корни.
Европа. Древнейшие достоверные следы культурной деятельности европейского человека относятся ко времени примерно 1 млн. лет назад, а возможно, и нескольно более раннему: до 2-2.5 млн. лет для Центрального массива Франции (Bonifay et al., 1989). Об этом свидетельствуют немногочисленные эоплейстценновые стоянки из южных и центральных районов Европы. Они расположены преимущественно в горных областях , как например, грот Валоне близ Ниццы в приморских Альпах (Южная Франция), с примитивными орудиями, по-видимому, виллафыриканского возраста. Датировку около 700 000 лет имеют остатки жилища эпохи нижнего палеолта из Пржезлетице и нижнепалеолетическая стоянка Странска Скала в бывшей Чехословакии. Можно предположить, что эти люди, скорее всего проникавши на юг Европы из Северной Африки, еще не обосновались здесь постоянно, так и костные остатки древнейших Ноmо в Европе имеют кромерский возраст(0.5, максимально до 0.7 млн. лет). Судя по всему, человек впервые появился в Европе на границе эоплейстоцена и нижнего плейстоцена.
В среднеплейстоценовое время – примерноот 0.4 до 0.2 млн. лет назад – гоминиды уже широко расселились по континенту: их ископаемые остатки обнаружены в Западной, Центральной, Южной и Восточной Европе. Наиболее ранними находками являются : Гейдельберг, или Мауэр (Германия), Араго и Монтморен (Франция), Вертешсёллеш (Венгрия), Петралона (Греция), Бильцингслебен (Германия), Атапуэрка (Испания) и др. Большинство среднеплейстоценовых людей Европы характеризуются сочетанием в разных пропорциях прогрессивных (сапиентных) и архаических черт и представляют собой “промежуточный” гетерогенный пласт между Н. Еrectus и Н. Sapiens. Так, человек из Вертешсёллеш, остатки которого вместе с чопперами обнаружены в травертиновом карьере недалеко от Будепешта, имел уже вполне современную абсолютную и относительную массу мозга. Емкость черепа у этого гоминида, жившего не позднее 350 000 лет назад, равна 1350- 1400 см. куб., в тоже время затылочная кость очень толста и массивна, с сильным наружным рельефом. В целом по форме черепа это гоминид ближе к прогрессивным “сапиентным” палеонатропам типа Схул, чем к типичным эректусам (синантропу) (В.В. Бунак). Таксономический диагноз ранних плейстоценовых гоминид Европы колеблется: поздний эректус, архаический сапиенс (пресапиенс), ранний неандерталец (пренеандерталец). Возможно, что популяции Н. Еrectus и Н. Sapiens вообще не были широко разделены во времени.
Дальнейший этап сапиентации представлен группой европейских палеоантропов древностью примерно 250 000- 200 000 лет. Сюда относятся прежде всего находки из Сванскомба (Южная Англия, близ Грейвсенда) и Штейнгейма ( Западная Германия, недалеко от Штутгарта). С этими гоминидами связана концепция “пресапиенса”, которая разрабатывается уже длительное время преимущественно зарубежными антрополагами. Предполагалось, что в эту эпоху уже можно морфологически отграничить линию, идущую к сапиенсу (“пресапиенсы”), от тупиковых ветвей предков неандертальцев (“пренеандертальцы”). Однако полученные дополнительные данные , расширившие представления о размахе изменчивости в популяциях гоминид, не подтвердили эту гипотезу. Возможно, что в верхнем и среднем плейстоцене Европы существовала единая сложная эволюционная линия или скорее пласт, характеризующийся постепенным накоплением сапиентных признаков в различных комбинациях с архаическими. Не все составляющие этого пласта достигли уровня сапиенса, однако по результатам анализа среднеплейстоценовых находок пока не удается верно выделить линию сапиенса. Отчасти это связано, конечно, с неполнотой и фрагментарностью многих материалов, как и с трудностями их точного датирования, но картину сапиентации, несомненно усложняла и возможность межпопуляционных контактов, создавшая основу для эволюции по “сетевидному типу” при ослаблении тенденций дивергентного видообразования. Чешский антрополог Я. Елик (1986) придерживается даже мнения о подвидовом ранге различий между Н. Еrectus, H. neanderthalensis и Н. Sapiens из-за мозаичности эволюции, которая не позволяет четко разграничить разные стадиальные группы гоминид (и тем более локальные варианты в пределах одной фазы).
Морфологическое разнообразие при разном соотношении архаических и сапиентных черт характерно и для европейских гоминид верхнеплейстоценового времени (Эрингсдорф, Саккопасторе, Фонтешевад и др.). Особого внимания заслуживает французская находка Фонтешвальда 1 древностью не менее 100 000 лет. На этом фрагменте черепа отсутствовал сплошной надглазничный валик, являвшийся почти облигатной принадлежностью “формирующихся людей”, в отличии от “готового человека”- сапиенса. Но находки из Фонтешвальда весьма неполные. Поэтому хотя и не исключена их принадлежность к группе архаического сапиенса, нельзя утверждать это с полной уверенностью.
Этап антропогенеза, непосредственно предшествовавший массовому появлению Н. Sapiens в Старом Свете, приходится на раннюю фазу последнего (вюрмского) оледенения. В это время в Европе широко распространилась основная культура среднего палеолита- мустье ( по названию грота Ле Мустье во Франции). Мустье- развитая каменная культура, характеризующаяся мультифункциональностью, представлена несколькими десятками типов орудий устойчивой формы. Среди них разнообразные остроконечники, скребла, скобели и др. В одной только Франции выделяют свыше 60 типов таких орудий (Борд). Для эпохи среднего палеолита характерно изготовление орудий на пластинах и отщепах, нередко с предварительной обработкой каменных заготовок - нуклеусов (техника леваллуа), вторичная обработка отщепов. Возможно появление в позднем мустье составных орудий (деревянного копья с кремневым наконечником); в некоторых мустьерских стоянках встречаются орудия из кости (острия, шилья, ретушеры).
Мустьерцы охотились на крупных животных (мамонта, пещерного медведя, носорога, сайгу, дикого осла, горного козла и т. д.), жили небольшими группами пл 20-30 человек. Социальная жизнь значительно усложнилась; некоторые археологи считают, что уже в эту эпоху начиналось становление родового общества, формирование определенных ритуалов (например, культ пещерного медведя в швейцарской пещере Драхенлох); проявляются черты заботы о соплеменниках (палеоантропы - инвалиды, дожившие до пожилого возраста); прослеживаются зачатки искусства, достигшего бурного развития в верхнем палеолите. Стоянки мустьерских людей разнообразны: открытые и пещерные, кратковременные охотничьи лагеря и долговременные базовые сооружения, мастерские. В мустьерскую эпоху в Европе и некоторых других районах Старого Света появились первые несомненные сапиенсы.
Наиболее распространенным вариантом гоминид в среднем палеолите Европы был неандертальский- прежде всего “классические” (или “типичные”), преимущественно западноевропейские неандертальцы.
Термин “неандертальцы” происходит от названия одного из первых местонахождений европейского плейстоценового человека, открытого в 1856 г. в Неандертале близ Дюссельдофа (Германия). Вид H. Neanderthalensis King выделен еще в 1864 г., но сейчас многие специалисты считают его не самостоятельным видом, а подвидом таксона Н. Sapiens- Н. Sapiens Neanderthalensis Основанием для такого объединения послужили новейшие исследования и результаты реконструкции физического облика, локомоции, образа жизни, интеллекта, речевых способностей, культуры неандертальцев, которые показали высокий уровень развития этих ископаемых людей. Другой аргумент- факты, свидетельствующие о возможном существовании неандертальцев и ранних “настоящих” сапиенсов (Н. Sapiens Sapiens) и смешениях между ними. Жесткая схема облигатной ассоциации физического типа неандертальцев с мустье, а сапиенса- с верхним палеолитом тоже полностью не подтвердилась.
Вюрмские неандертальцы населяли преимущественно приледниковую зону Европы: большая часть находок сделана южнее 50 градусов с. ш. , по которому проходила граница максимального (рисского) ледника и зоны вечной мерзлоты. Особенно много остатков ископаемых людей известно с территорий Франции, Бельгии, Германии, Италии, Испании, Бывших Югославии и Чехословакии. На территории бывшего СССР ископаемые люди неандертальского типа обнаружены главным образом в Крыму. Но классические неандертальцы жили не только в Европе; известны их находки в Северной Африке (Марокко), Передней и Средней Азии, возможно, и в других районах Земли. Хронологические границы существования классических неандертальцев: около 70 000-35 000 лет назад.
Физический тип гоминид имел отпечаток адаптации к суровым условиям перигляционной зоны: широкоплечесть при сравнительной малорослости (длина тела мужчин в среднем 160-163 см.), некоторое укорочение “рефрижераторных органов” (предплечья, по сравнению с плечом и голени сравнительно с бедром), высокое отношение массы тела к его поверхности, сильное развитие мускулатуры и костяка, внутренняя массивность скелета (высокий уровень компактизации), которая могла быть следствием функциональной адаптации или проявлением специфических особенностей обменно-гормонального статуса, но в далеко зашедших случаях, возможно, вызывала вторичную анемию из-за атрофии гемопоэтической такни. Все эти признаки, особенно уменьшение поверхности тела по отношению к его объему (т. е. Теплоотдачи), рассматриваются в качестве адаптаций к холодному или умеренному климату начала последнего оледенения. Грубость и массивность костяка объясняется также необходимостью противостоять различным силовым механическим нагрузкам. Эта особенность проявлялась уже в раннем детстве. У 6-8 месячного ребенка неандертальца из Крыма (Киик-Коба) тип поперечного сечения ребер массивный, округленный, а не в виде вытянутого овала , как у современного человека. Исключительно толстостенность костей конечностей и даже ключиц у 8-9 летнего неандертальца из Узбекистана (Тешик-Таш). По-видимому, продолжительность жизни неандертальцев была невелика, и они крайне редко доживали до конца репродуктивного периода.
Мозг неандертальцев крупный, его объем нередко превышал средний для современного человека; по реконструкциям эндокранов предполагается праворукость. Череп характеризуется большими размерами, удлиненной формой (долихокранией), массивным надглазничным торусом, низким сводом и скошенным лбом. Затылок обычно в форме “шиньона”. Такое удлинение задней части черепа рассматривают как “противовес” очень крупному, длинному, высокому и массивному лицевому отделу с большими, часто очень стертыми передними зубами, вздутой верхней челюстью, отсутствием клыковых ямок. Эти своеобразные черты связываются главным образом с адаптацией к мощным силовым нагрузкам со стороны челюстного аппарата, особенно латерально расположенных клыков.
Типичная неандертальская кисть отличается от современной исключительной массивностью трубчатых костей, расширением запястья, нестабильностью формы сустава 1 луча, наличием компенсаторных признаков, облегчающих противопоставление большого пальца. Иными словами, многие черты кисти отражают адаптацию к силовому захвату, что, возможно, делало руку неандертальца менее ловкой по сравнению с ее современным типом.
Все эти особенности, разумеется, придавали облику неандертальцев определенное своеобразие, но не отделяя их глухой стеной от остальных гоминид. Почти все перечисленные признаки, особенно в смягченном виде, были достаточно распространены среди палеоантропов, встречались они и раньше и позднее. Поэтому состав неандертальского вида (или подвида) может быть более широким, с включением в него не только поздних вюрмских неандертальцев, но и многих более ранних палеоантропов или даже всех палеоантропов вообще.
Первые мустьерские архаические сапиенсы Европы известны из некоторых ее южных и восточных районов: Крыма (Староселье), Приазовья (Рожок), Кавказа (Ахштырская пещера между Сочи и Адлером), возможно, бассейна Днепра (Романково, Самара). Особенно большое значение имеет находка неполного скелета ребенка 1.5-2 лет из пещеры Староселье (близ Бахчисарая) на долговременной охотничьей стоянке. У этого гоминида преобладают черты сапиенса в сочетании с культурой позднего мустье. Чрезвычайно пеструю и даже загадочную картину представляют находки из пещеры Крапина близ Загреба (бывшая Югославия) древностью от 80-70 до 30 тыс. лет назад. Здесь наряду с неандертальцами обнаружены и остатки людей более сапиентного типа- с прямым, довольно грацильным лбом, выступающим подбородком и т. д. В Центральной Европе прослеживается определенная морфологическая преемственность с усилением признаков сапиентации между сравнительно ранними палеоантропами, более поздними их формами и далее- к ранним неоантропам. “Переходный характер имеет, видимо, и череп мустьерского ребенка из пещеры Пеш-де-Лазе (Франция).
Азия. Пока нельзя сказать определенно, когда появились древнейшие гоминиды на азиатском континенте. Выше упоминалось мнение, что среди плиоценовых и раннеплейстоценовых гоминидов- гигантопитеков и мегантропов- были и представители азиатской ветви массивных австралопитеков. Высказывалось также предположение, что некоторые азиатские находки гоминид, например, ребенок из Моджокерто (Ява), могут быть причислены к виду H habilis. Однако малый возраст, фрагментарность находки и, главное, невысокая древность (наиболее вероятная дата 0.6-0.7 млн. лет) не позволяют сделать столь определенный вывод об ее таксономическом положении. В целом же хронология ранних гоминид Юго-Восточной Азии (0.8- 0.5 млн. лет для большей части находок индонезийских питекантропов) соответствует временным границам эректусов, но не хабилисов.
Стоянки архантропов в Индокитае (Таиланд) также имеют древность 0.7-0.6 млн. лет назад.
Если судить по археологическим материалам, первые гоминиды могли проникнуть в Азию еще в плейтоцене: известна датировка 2 млн. лет для раннешальской стоянки Убейдиа (Израиль). Но физический тип носителей этой культуры остается пока неизвестным. В ашельскую эпоху человек уже заселил все основные районы Азии- Переденей, Среденей, Южной, Юго-Восточной и Восточной. Существование в Азии древнейшей олдувайской культуры остается под вопросом. Хотя галечные орудия были обнаружены в нескольких местонахождениях на Адлтае и на Дальнем Востоке, по-видимому, также в Якутии, имеются расхождения в их датировании. Так, для нижнего слоя стоянки на р. Улалинке (район Горно-Алтайская) датировки колеблются от верхнего плиоцена до нижнего плейстоцена и даже позже. Известно, что в некоторых регионах Азии галечная культура существовала и в среднем плейстоцене.
В Восточной и Юго-Восточной Азии хронологическая и морфологическая преемственность гоминид более отчетлива на ранних этапах их эволюции. В Индонезии она очевидна для питекантропов Явы (1.2-0.5 млн.лет) и “явантропов” из Нгандонга (солосский человек древностью около 0.1 млн. лет назад). При сравнении этих групп гоминид прослеживается небольшое увеличение объема мозга: в среднем от 930 до 1151 см куб. ; общей чертой является массивный череп со слабоизогнутым основанием. Значительно сложнее вопрос о преемственности на более поздних фазах сапинтации. Иногда ее пытаются установить по линии яванских гоминид: поздний солосский питекантроп (Нгандок)- сапиенс без определенной датировки (Ваджак), эта форма, впрочем, достаточно атипичная и неконсолидированная. Довольно гипотетична и другая схема реконструкции: поздние питекантропы Явы- позднеплейстоценовый сапиенс из Австралии (Кау-Свэмп). Древнейший датированный сапиенс австрало-азиатского региона- юношеский череп протоавстралоидного типа из пещеры Ниа на Калимантане, имеющий древность около 40 000 лет. Высказывается гипотеза, что заселение Австралии в верхнем палеолите происходило популяциями двух типов: более массивные гоминиды проникали на запад континента с Явы и из соседних областей, а более грацильные- на северо-восток; возможно, они были связаны в своем генезисе с Южным Китаем и мигрировала в Австралию через Индокитай, Калимантан, Новую Гвинею.
В восточноазиатском регионе (Китай) хронологическая и морфологическая преемственность снова более определена на уровне ранних- гоминид: от древнейшего (около 1 млн. лет) эректуса из Данау (провинция Юньнань) и примитивного синантропа латьянского (около 0.7 млн. лет) с малой емкостью черепа (775-7823 см. куб.)до более позднего и прогрессивного синантропа пекинского древностью от 460 000 до 230 000 лет назад с более крупным мозгом (в среднем 1040 см. куб.) Сходную древность имеют и остатки Н. Еrectus из Вьетнама (провинция Лангшон на границе с КНР): 300 000- 250 000 лет.
Далее можно упомянуть некоторые находки, обнаруживающие в морфологическом отношении переходные черты от синантропа к сапиен-су. Это предполагаемые более прогрессивные потомки синантропа, остатки которых обнаружены близ деревни Суйцзияо, или раннесапиентный человек из Дали (пров. Шэньси) древностью около 100000 лет. Китайские антропологи и археологи указывают на существование в местонахождениях некоторых архаических сапиенсов, например человека из Динцуна с предположительным возрастом 90 000-70 000лет, преемственности и археологических традиций с чжоукоудяньской культурой синантропа. Впрочем, не исключено и более раннее возникновение архаического Н. sapiens с признаками Н. erectus в Восточной Азии, если судить по недавней (1984 год) находке черепа на стоянке Цзиньню-шань в провинции Ляонин (северо-восток КНР) с датировкой не менее 0,2 млн лет (урановый метод).
К группе “переходных вариантов”, или ранних сапиенсов, относятся также Мапа, Чяньян, Ордос (Внутренняя Монголия). По большей части эти материалы фрагментарны и слабо датированы, но, принимая во внимание и археологические данные, нельзя исключить вероятность преемственного развития гоминид и на восточной окраине ойкумены. Черты сходства с синантропом отмечаются и для одного из ранних сапиенсов из пещеры Минатогава в Японии (18 000 лет). В то же время обращает на себя внимание большое морфологическое разнообразие и отсутствие специальных черт сходства с синантропами из Чжоукоудяня у обнаруженных в непосредственной близости о этого местонахождения верхнепалеолитических людей из так называемой шандмидунской популяции. В Передней Азии в палеолите были благоприятные условия для существования гоминид. Здесь сделаны важные находки “сапиентных мустьерцев” Многие специалистым относят к числу ранних сапиенсов мустьерского человека из пещеры Кафзек с датировкой 92 000+-5 000 лет (термолюминесцентный анализ) или 115 000+- 15 000 лет (урановый метод), а также наиболее прогрессивные формы гоминид из пещеры Схул (близ г. Хайфы)- Схул IV и Схул V. Все эти мустьерцы имеют признаки сходства с кромпаньонцами- наиболее известными представителями неоантропов из верхнего палеолита Европы и некоторых других регионов. Среди них были высокорослые длинноногие люди с пропорциями конечностей (удлинение голени относительно бедра и предплечья относительно плеча), типичными для современных тропических групп- австралийских, негрских и др. У средне- и верхнепалеолитического человека такие “тропические” пропорции, видимо, ассоциировались с большой скоростью бега, ловкостью в прыжках и метании предметов, наблюдающейся и у современных длинноногих групп. Однако среди палестинских гоминид были и неандертальцы, как Схул IX, Табун 1, Кебара. Другие переднеазиатские и среднеазиатские люди (Шанидар из Ирака, Амуд из Израиля, Тешик-Таш из Узбекистана) тоже обнаруживают переходные к сапиенсу черты при значительной еще “неандерталоидности” в структуре надглазничного рельефа, выступании средней части лица и других признаках. Возможно, что недавняя находка остатков скелета ребенка из ашельской пещерной стоянки Сельунгур в Средней Азии (Узбекистан) принадлежит к кругу эректуса.
Плейстоценовые гоминиды на территории бывшего СССР
В эоплейстоцене не обнаружено абсолютно достоверных следов деятельности ископаемого человека на территории нашей страны. Наиболее вероятную эоплейстоценовую датировку имеет галечная культура из многослойной пещеры Азых (Нагорный Карабах).Имеются также сведения об остатках галечных оле- культур не вполне ясной датировки в районе Алтая (Улалянка), на Дальнем Востоке (Филимошки, Усть-Ту, Кумары), в Якутии (Диринг-Юрях) и др. Предполагаемый возраст этих стоянок колеблется от верхнего плиоцена до среднего плейстоцена (ввиду отсутствия четких геологических датировок ,и существования галечной техники в некоторых районах Сибири вплоть до неолита).
По-видимому, заселение территории бывшего СССР в основном происходило в среднем плейстоцене. По археологическим данным, возможны миграции палеолитического населения из нескольких центров: территория Русской равнины заселялась преимущественно из Южной и Центральной Европы, Кавказ — из Передней Азии, Крым — частично из Европы, частично из Передней Азии, Средняя Азия — тоже из нескольких центров (Передней и Юго-Восточной Азии). Схема эта пока довольно гипотетична.
География раннепалеолитических памятников свидетельствует о довольно широком распространении человека в среднем плейстоцене: они известны из районов Кавказа, Молдавии, Приднестровья, Приазовья, бассейна Днепра, Закарпатья, Средней Азии, Казахстана и других мест. Однако палеоантропологические материалы этой эпохи еще скудны (рис. 34). Большая их часть имеет неясную датировку, например нижняя челюсть так называемого “азыхантропа” (1968 г.) .из Азыхской пещеры на Кавказе, характеризующаяся переходными чертами от архантропа к палеоантропу и напоминающая некоторых европейских “пренеандертальцев” (Араго, Гейдельберг). В Ереване на левом борту каньона р. Раздан в верхней части аллювиальной террасы, предположительно датируемой ашелем, в 1975 г. обнаружен череп человека весьма сапиентного облика, но с пониженным сводом и толстыми костями; однако, геологическая ситуация в районе находки не вполне ясна, и ее датировка нуждается в уточнении.
В Узбекистане близ г. Ферганы известна палеолитическая многослойная пещерная стоянка Сельунгур, расположенная на высоте 2000 м над ур. моря. Открытые в III культурном (ашельском) слое зубы и фрагмент плечевой кости ребенка около 10 лет с очень высокой кортикализацией, возможно, принадлежат представителю локального варианта Н.erectus.
В конце среднего плейстоцена на территории Русской равнины началось распространение максимального днепровского (рисского) оледенения. В то время ледники доходили до уровня современного Днепропетровска. Достоверно датированных находок человека днепровской эпохи не обнаружено.
В начале верхнего плейстоцена (рисс-вюрмское, или микулинское, межледниковье) климатические условия становятся более благоприятными. Значительно возрастает освоенная человеком территория. В эпоху мустье гоминиды обитали и в более северных районах—до 52° с. ш. в бассейнах Десны и Волги, но большая часть стоянок сосредоточена на юге: Крым, Кавказ, Русская равнина, Средняя Азия, Казахстан. Особенно много мустьерских памятников в Крыму, бассейне Днестра и Прута, в Приазовье, Поднепровье. Геологически начало эпохи мустье скорее всего соответствует никулинскому интергляциалу; нет достоверно датированных мустьерских стоянок домикулинского времени.
В принципе допускается возможность автохтонного развития в некоторых районах мустьерских культур на основе местного ашеля, как, например, на Русской равнине или в Закарпатье (стоянка Королеве с “переходными” чертами).
Наиболее заселенным районом в мустьерскую эпоху был Крым, где обнаружено свыше 30 стоянок. Отсюда же известны и самые полные палеоантропологические материалы. Основные находки сделаны: в пещере Киик-Коба близ Симферополя (1924 г.), где открыты 2 'неполных скелета неандертальцев—взрослого и ребенка 6—8 мес; на стоянках Заскальная V и VI недалеко от г. Белогорска (1969—1973 гг.) —3 индивида неандертальского типа, из которых один взрослый и двое детей в возрасте примерно 5 и '10—12 лет; в поселке Староселье близ г. Бахчисарая (1953 г.) —неполный скелет ребенка сапиенса 1,5—2 лет. Примечательно, что все это хотя и различающиеся в хронологическом отношении, но однокультурные памятники. Прослеживается археологическая преемственность более ранних (Киик-Коба, видимо, Заскальная, относящаяся к концу микулинского времени) и поздних (Староселье— первая половина валдая) мустьерских стоянок.
Несколько находок мустьерского человека известны из Приазовья, Прикубанья и Закавказья. Это датирующаяся, по-видимому, микулинским временем многослойная стоянка Рожок, 1 на .северном побережье Таганрогского залива, в IV слое которой обнаружен верхний моляр с преобладанием черт сапиентного типа (1961-1962 гг.); Ахштырская пещера между Сочи и Адлером (1961 г.) с очень фрагментарными остатками (моляр и 3 плюсневые кости) человека сапиентного типа; Лжручула (1960 г.) и Сакажиа (1974 г.) в Западной Грузии с фрагментарными находками палеоантропов, имеющих признаки сходства с переднеазиатскими мустьерцами; нижняя челюсть неандертальского ребенка 2—3 лет без подбородочного выступа, но с некоторыми сапиентыми чертами из Баракаевской стоянки в Прикубанье (Западный Кавказ 1979 г.).
В бассейне Днепра известны остатки человека сапиентного типа из нескольких переотложенных мустьерских стоянок без отчетливой геологической датировки. К их числу относятся: Романково (1957 г.)— сильно фоссилизированная бедренная кость и 2 плечевые кости высокорослого человека с преобладанием черт сапиенса; Самара (левый приток Днепра) — находка в размытом мустьерском слое фрагмента очень минерализованной плечевой кости, принадлежавшей высокорослому сапиенсу с примесью архаических особенностей; фрагменты трех черепов сапиенса с неандертальскими признаками из района Днепропетровска (1947—1948 гг.). Синхронность всех этих остатков с мустьерскими орудиями вполне возможна, хотя и не бесспорна. Археологами отмечаются черты сходства между орудиями, обнаруженными в Романкове (двусторонние треугольные наконечники) и в стоянках Крыма, например в Староселье. Несмотря на неясный геологический возраст некоторых находок появление человека сапиентного типа с пережиточными архаическими чертами в позднем мустье Крыма и Русской равнины представляется весьма вероятным. К этому кругу находок, по-видимому, можно отнести и фоссилизированную лопатку из местонахождения Шкурлат III (Воронежская обл.), датируемого микулинским межледниковьем. Она принадлежала высокорослому человеку современного типа с некоторыми архаическими признаками. Весьма многочисленны мустьерские стоянки в Казахстане и Средней Азии; мустьерцы обитали также в Сибири, на Алтае и Дальнем Востоке.
Единственная пока находка человека мустьерскои эпохи в Азиатской части бывшего СССР—ребенок палеоантропа 8—9 лет из пещеры Тешик-Таш в Южном Узбекистане (1938 г.). Неполный скелет тешикташца обнаруживает комбинацию преобладающих неандерталёских и сапиентных признаков; к числу последних относятся, например, переходные черты в строении эндокрана и кинодонтия (зубы с узкой полостью). Эта находка показывает, что Средняя Азия также могла входить в территорию сапиентации.
В верхнем палеолите происходило дальнейшее расширение обитаемой зоны—до 71° с. ш. Она уже включала бассейны Енисея, Алдана, Лены, Индигирки. Нет единого мнения о хронологическом рубеже верхнего палеолита. Скорее всего на Русской равнине он начался во время молого-шекснинского межледниковья. Этот период включает и последующее, самое сильное осташковское похолодание, когда ареал обитания человека сократился. Судя по некоторым археологическим и палеоантропологическим данным, возможно развитие человека верхнего палеолита на юге Русской равнины на основе местного мустьерского населения (преемственность археологических культур в Костенках и отчасти в Сунгире со Старосельем, постепенное усиление признаков сапиентации от более ранних к более поздним мустьерским популяциям: Киик-Коба и Заскальная, с одной стороны, и Староселье и Рожок—с другой). Палеоантропологические материалы верхнего палеолита наиболее многочисленны в европейской части бывшего СССР. Это прежде всего стоянки бассейнов Дона и Клязьмы — Костенки и Сунгирь. На Кавказе и в Средней Азии остатки гоминид этой эпохи малочисленны.
Костенки (1952—1954 гг.)—крупная верхнепалеолитическая стоянка—поселение в долине Дона (Воронежская обл.). Ее абсолютный возраст около
30 000—25 000 лет назад. Морфологически население стоянки разнообразно: Костенки II—взрослый мужчина кроманьонского типа; Костенки XVIII—ребенок 9—II лет, сближающийся с ним по типу; Костенки XIV (Маркина Гора)—наиболее полный и ранний по времени скелет современного человека с некоторыми чертами экваториального типа (пропорции конечностей, очень низкое отношение массы тела к поверхности, прогнатизм, широкий, хотя и сильно выступающий нос); Костенки XV—ребенок 5—6 лет, имеющий признаки сходства с центральноевропейскими находками, например из Пшедмо-сти ( бывшая Чехословакия).
Стоянка Сунгирь (1956—1977 гг.) расположена на окраине г. Владимира в бассейне Клязьмы; она относится к концу мологошекснинского межледниковья, абсолютный возраст 25 000—24 000 лет назад. Обнаружены остатки всего 8 индивидов, из них наиболее полные: взрослый мужчина Сунгирь 1, дети Сунгирь II (II—13 лет) и Сунгирь III (9—II лет). Население стоянки морфологически разнообразно; таксономически это сапиенс с кроманьоидными и некоторыми более архаическими, в том числе неандертальскими, признаками. Характерны: высокорослость, большая ширина плеч, удлинение средних отделов конечностей, укрепление запястья (макрокарпия), “саблевидная голень” и др. Черты сходства отмечаются с “кроманьоидными мустьерцами” из Передней Азии типа Схул и центральноевропейскими неоантропами из Пшедмости. Археологический инвентарь стоянки—верхний палеолит с некоторыми мустьерскими категориями, а также богатым набором костяных орудий и украшений, копьями из выпрямленных бивней мамонта. Прослеживается генетическая связь с памятниками костенковско-стрелецкой культуры на Дону.
В азиатской части СССР палеоантропологические находки этого времени очень скудны. Условно к верхнему палеолиту относят фрагментарные черепа из Самаркандской стоянки в Узбекистане (1962, 1966 гг.), возможно, кроманьоидного типа, и находку Афонтова Гора “близ Красноярска (1937 г.) с абсолютной датировкой 20900 лет— фрагмент черепа сапиенса с уплощенным переносьем; известно также погребение ребенка плохой сохранности на стоянке Мальта (бассейн Ангары).
Полученные на основе археологических и палеоантропологических материалов данные позволяют предположить, что территория Восточной Европы входила в зону сапиентации, причем сапиенс впервые появился здесь в эпоху мустье. Обращает на себя внимание и некоторое сходство обнаруженных в мустьерских и верхнепалеолитических стоянках гоминид (Романково, Самара, Сунгирь, Пшедмости и др.). На этих территориях известны и более поздние находки “переходных форм” с неандерталоидным налетом и не вполне ясной датировкой: черепные крышки из районов Северного Кавказа (Подкумок, 1918 г.), бассейна Волги (Хвалынск, 1927 г.), Подмосковья (Сходня, 1936 г.).
Северо-восток Русской равнины заселялся в молого-шекснинское время; открыты первоклассные верхнепалеолитические памятники на восточной окраине Европы, как, например, Каповая пещера на Урале или Бызовая стоянка на Печоре, датируемая 25 450 лет назад. Освоение северо-западных районов европейской части бывшего СССР, включая бывшую Прибалтику и Белоруссию, протекало, видимо, уже в позднеледниковое время, на рубеже плейстоцена и голоцена. Постепенное расширение ареала в северном направлении происходило и в азиатской части бывшего СССР. В мезолите человек проник в районы, расположенные севернее 72° с. ш. (Таймыр, Заполярье), чем в основном и было завершено первоначальное заселение территории СССР.
В заключение всего этого краткого обзора палеоантропологических материалов, относящихся к “переходной эпохе”, можно сделать вывод, что процессы сапиентации в Старом Свете не ограничивались каким-то одним регионом, но протекали на разных территориях и на основе достаточно разнообразного морфологического субстрата.
Время и место возникновения Ноmо sapiens. Гипотезы моно- и полицентризма
Заключительный этап сапиентации занял широкий хронологический интервал: от 0,35—0,25 до 0,04—0,03 млн лет назад. Пока точно неизвестно, происходил ли этот процесс путем кладогенеза, т. е. ветвления линий, способом филогенетической дифференцировки или же путем анагенеза—постепенным изменением одного таксона в другой при общем подъеме уровня организации, хотя, как упоминалось выше, более вероятной представляется вторая гипотеза. Высказывается и предположение, что могло иметь место сочетание обоих этих способов эволюционного развития, и ветвление в пределах целостных полиморфных групп дополнялось “прямым” развитием отдельных ветвей (В. В. Бунак).
В науке обсуждаются несколько возможных уровней выделения линии сапиенса: от эоплейстоцена до вюрма. В качестве примера гипотез ранней дивергенции можно назвать концепцию фамногенеза (от греч. “фамнос”—куст) В. В. Бунака (1980), который признает исключительную древность филогенетических корней сапиенса, выделяя его “ветвь” в эоплейстоцене от австралопитекоидного (или даже от еще более раннего) предка. Эта схема обосновывается главным образом находками ранних прогрессивных гоминид в Восточной Африке, как, например, уже упоминавшейся выше “челюсти ЕR 730” из Кении. Она близка к взглядам Г. Хеберера (1944 г. и далее), отраженным в его “гипотезе развертывания”: дивергенция линий сапиенса без сплошного надглазничного торуса (“апроскопины”) и линий архантропов и палеоантропов с сильным рельефом черепа (“проскопины)” от австралопитекоидного предка с умеренно развитым торусом (“олигопроскопины”). В данном случае в основу филогенетической реконструкции положен сложный и достаточно изменчивый признак неясного генезиса — рельеф лобной кости, значение которого тем самым неоправданно преувеличено. Кроме того, если судить по высокому, т. е. типичному для регрессивных структур скелета, уровню изменчивости выступания надглазничного” рельефа у современного человека, в числе его ближайших предков вряд ли были гоминиды с очень слабым развитием или даже отсутствием торуса, как считал Хеберер. В целом гипотезы ранней дивергенции, исключающие из эволюции сапиенса не только неандертальцев, но и эректусов, пока мало обоснованы.
Наиболее распространено мнение о средне- или ранневерхнеплей стоценовом времени возникновения линии сапиенса. В качестве его вероятных предков в разных вариантах филогенетических схем фигурируют различные гоминиды этой эпохи: Вертешсёллеш, Сванскомб и другие находки. Однако выше уже отмечалось, что пока нет возможности достаточно четко дифференцировать линии “пресапиенса” и “пренеандертальца” в среднем плейстоцене. Это наглядно иллюстрируется судьбой весьма распространенной в прошлом концепции “пресапиенса”, согласно которой в среднем плейстоцене уже произошла дивергенция обеих линий: “пресапиенса”, представленной находками Сванскомб и, далее Фонтешевад, и “пренеандертальца”, в основе которой череп из Штейнгейма (Г. Хеберер, А. Валлуа, А. Тома и др.). Однако в последующем обе находки были отнесены к единому кругу ископаемых гоми-кид. Столь же зыбкими могут оказаться и другие, предложенные позже, варианты этих прототипов. Среди более современных модификаций гипотезы пресапиенса можно привести в качестве примера упоминавшуюся выше схему В. В. Бунака (1980), в которой в роли “пресапиенсов” фигурируют уже Сванскомб и Штейнгейм вместе, а “пренеандертальца-ми” являются Араго и Монтморен, или схему Дж. Хайм (1987), согласно которой от архаических плейстоценовых гоминид (Араго, Мауэр) дивергируют линии собственно “пресапиенса” (Фонтешевад) и “пренеандертальца” (Петралона).
Выше рассмотрены некоторые кладогенетические гипотезы сапиентации. Однако пока нет прямых доказательств, что процесс сапиентации осуществлялся путем кладогенеза. Если же придерживаться концепции анагенеза (ароморфной эволюции), более отвечающей специфике поздних этапов антропогенеза, то общая картина сапиентации предстает как неравномерная мозаичная эволюция различных групп палеоантропов, сопровождавшаяся смешениями, потоками генов между популяциями, замедлявшими их дивергенцию и приводившими к развитию по сетевидному типу. При таком способе эволюционного развития было бы, конечно, крайно сложно выделить конкретных “предков”.
По-видимому, отдельные признаки сапиенса могли появляться очень рано, еще даже в эоплейстоцене. Морфологическая изменчивость гоминид вообще велика, и, вероятно, немаловажную роль играли здесь и повторные мутации. Напомним, что подбородочное возвышение известно уже на нижней челюсти восточноафриканского человека из Кении, жившего около 2 млн лет назад, тогда же отмечено и довольно грацильное строение лба; редукция зубов мудрости отчетлива на челюсти примитивного лантьяньского синантропа. Подобные сапиентные тенденции могли возникать в отдельных группах гоминид давно и неоднократно, и в этом смысле можно сказать, что сапиентация имеет глубокие корни, хотя древнейшие “настоящие сапиенсы”, видимо, пока неизвестны ранее 0,1, максимально—0,2—0,25 млн лет назад. Объединение Н.sapiens и Н. пеапderthalensis в рамках одного вида естественно также может удревнить возраст таксона. Последнее внесло значительные изменения в концепцию так называемой “неандертальской фазы”, впервые предложенной А. Хрдличкой более полувека назад. Речь шла о возможности происхождения сапиенса от классических неандертальцев вюрьма.
.Роль поздних неандертальцев в эволюции человека уже длительное время остается предметом дискуссий. Если таксой Н. Erectus обычно рассматривается как облигатное звено филогенеза Н. sapiens, то концепция “неандертальской фазы” не является общепризнанной. Крайняя точка зрения—исключение вюрмских неандертальцев вообще из числа прямых предков неоантропа (гипотеза “пресапиенса”) или допущение такой возможности в ограниченной степени и для внеевропейских территорий (например, в Азии от неандертальцев типа Шанидар и Тешик-Таш). При этом принимается возникновение европейского сапиенса от ранних пресапиенсов Европы либо более поздняя миграция сапиенса африканского происхождения в Европу из Африки и (или) Азии.
Сторонники происхождения Н. Sapiens от местных неандертальцев Европы указывают на существование морфологической преемственности между ними в некоторых районах, как, например, в Центральной Европе. Такова реконструируемая по хронологическому и морфологическому принципам линия: ранние неандертальцы (Крапина, Гановцы, Шубаюк, Охос)—поздние неандертальцы (Виндижа, Шипка, Шаля)—ранние сапиенсы (Ветерница, Пшедмости, Брно, Долни-Вестонице и особенно Младеч). В пользу этого взгляда приводятся и факты из области археологии—преемственность позднемустьерской культуры классических неандертальцев и верхнепалеолитической индустрии сапиенса. Указывается также, что если составить комбинированную популяцию из неандертальцев и сапиенсов Европы и Ближнего Востока, то, по краниологическим данным, она обнаруживает не большее отклонение от нормального распределения, чем каждая из этих двух групп в отдельности. Отсюда заключают, что эволюционная трансформация черепа могла реализоваться без существенного нарушения генетического равновесия.
К этому можно добавить, что “переходные формы” встречаются во многих районах Европы в эту эпоху и позднее (Подкумок, Хвалынск, Сходня); неандерталоидные черты отмечаются у многих мустьерских гоминид Восточной Европы и Крыма. По-видимому, в каких-то случаях нельзя исключить возможность филетической преемственности неандертальцев и сапиенсов, тогда как в других “переходные формы” могли быть результатом их поздних смешений. Так, по имеющимся данным, в Передней Азии не прослеживается филетическая преемственность неандертальцев и неоантропов, так как архаический сапиенс (Кафзех) на несколько десятков тысяч лет древнее неандертальцев (Кебара, Табун 1).
Если признать—в соответствии с изменившимися представлениями о таксономическом положении неандертальского человека—достаточно широкую хронологическую и территориальную репрезентативность таксона Н. пеапderthalensis, то неандертальская фаза становится, очевидно, закономерным этапом на пути сапиентации. Если же придерживаться мнения о более узких рамках этого таксона, то неандертальцы могли являться предками сапиенса наряду с другими ископаемыми гоминидами (поздними прогрессивными эректусами или пресапиенсами). Пока остается открытым вопрос: произошло ли около 40000 лет назад вытеснение неандертальцев сапиенсом (с метисацией или без нее), или их эволюционная трансформация в неоантропов (полная или частичная). Одна из недавно высказанных гипотез трансформации (Василик и др., 1989) связывает ее с процессами акселерации, приведшими к значительному изменению физического типа в ходе сапиентации, возможно, в результате воздействия такого глобального фактора, как напряженность геомагнитного поля Земли. схемой
В науке существуют две основные гипотезы о числе центров сапиентации и их локализации на земном шаре: это гипотезы моно- и полицентризма. Разногласие между ними нередко выступает в форме дискуссии между сторонниками “миграционной” и “эволюционной” теорий (У. Хоуэлл).
В отечественной антропологии в течение многих лет разрабатывалась гипотеза “широкого моноцентризма” (Я. Я. Рогинский, 1947, 1949, 1969, 1972 и др.), т. е. происхождения Н. sapiens на одной, хотя и достаточно обширной, территории Средиземья (Восточная Африка, Передняя Азия, Южная и Юго-Восточная Европа). Другой вариант моноцентризма, получивший наибольшее распространение в современной зарубежной антропологии,—концепция африканской (афро-европейской) прародины. Происхожденние неоантропов связывается с Африкой, откуда “протокроманьонцы” мигрировали в Европу—непосредственно или-через переднюю Азию (П. Эндрюс, Г. Брейер, Р. Протш, Л. Шотт„ X. Стрингер и др.). Так, Л. Шотт (1985, 1986) считает, что предок, от которого произошли линии Н.sapiens и Н. Sapiens neanderthalensis, обладал мозаикой признаков—архаических и прогрессивных. Заселение Европы происходило из Северной Африки несколькими волнами. Классические неандертальцы в дальнейшем сформировались под влиянием изоляции и генного дрейфа, но более поздние мигранты Н. Sapiens sapiens смешивались с неандертальским населением, являвшимся потомками более ранних переселенцев. В соответствии со схемой П. Эндрюса (1984) только “западные” (африканские) эректусы являлись предками Н. sapiens. Выделяются две миграционные волны из Африки: в результате первой миграции возникли европейские неандертальцы, а второй—палеоантропы типа Схул-Кафзех и современный человек. С точки зрения африканской гипотезы можно было бы объяснить высокорослость и “тропические” пропорции некоторых ранних мустьерских сапиенсов Передней Азии и, по-видимому, Юго-Восточной Европы. Однако справедливость мнения об их экваториальном происхождении следует еще обосновать уточнением датировок средне- и верхнеплейстоценовых находок из Европы, Передней Азии и Африки, которые пока трудно сопоставлять.
В качестве аргументов моноцентризма используются и биомолекулярные данные. Так, на основе анализа генов митохондриальной ДНК у представителей различных современных рас А. Уилсон и др. (1982) выдвигают гипотезу о резком уменьшении численности гоминид в верхнем плейстоцене и возникновении современного человека около 200 000 лет назад в Африке южнее Сахары, откуда примерно 100 000 лет назад началось его расселение с последующей изоляцией и генным дрейфом. Данные современной популяционной генетики показывают высокую генетическую дифференциацию африканских популяций и, следовательно, длительное время их становления. Предполагается, таким образом, что все современное генетическое разнообразие человечества берет начало в восточноафриканском генетическом пуле.
Однако есть и мнение о происхождении сапиенса от “восточного” (азиатского) ствола Н.erectus (Варк, 1983). При этом речь идет о прямой миграции сапиенса из Азии или “обходной”—из Африки через Азию в Европу. Эта концепция обосновывается преимущественно краниологическими материалами.
Основателем гипотезы полицентризма, т. е. нескольких центров происхождения сапиенса, является американский антрополог Ф. Вейденрейх (1938, 1943). Им выделены 4 таких региона формирования человека современного типа и его рас: Юго-Восточная Азия (австралоиды),Южная Африка (капоиды или негроиды). Восточная Азия (монголоиды) и Передняя Азия (европеоиды). По своим теоретическим воззрениям Вейденрейх был механоламаркистом, признававшим изначальную способность организма к целесообразным реакциям; в основе его полицентризма лежал ортогенетический принцип. Большую глубину расовых различий признает и другой американский антрополог—К. Кун (1963), выделивший пять независимых центров трансформации Н. Erectus в H. sapiens
Среди современных зарубежных полицентристских схем одна из наиболее разработанных—гипотеза А. Тома (1962, 1964, 1973), в соответствии с которой выделяются три основных очага сапиентации: европеоидно-негроидный (западный), монголоидный и веддо-австрало-айно-идный (восточный). Допускается несколько миграционных волн из Африки в Европу в плейстоцене, в периоды обмеления Средиземного моря, соответствующие ледниковым эпохам. Древнейшая из них представлена популяциями типа Сванскомб-Фонтешевад с примитивной культурой отщепов, вторая—типа Штейнгейм с культурой рубил и третья, позднейшая,—типа Неандерталь с культурой мустье. В межледниковые периоды потомки этих переселенцев оставались в изоляции, причем каждая последующая волна оттесняла предыдущую на север и восток. Это приводило к дивергенции, в итоге которой вытесненные неандертальцами потомки популяций “пресапиенсов” (Сванскомб-Фонтешевад) положили начало формированию гетерогенной группы западных неоантропов, происходившему на территории Юго-Восточной Европы (в понто-кавказ-ской области); в дальнейшем эти популяции сапиенсов продвигались на запад—в Переднюю Азию и Европу, где они вытеснили неандертальцев и заняли их место. В Передней Азии эти неоантропы смешались с местными палеоантропами — неандертальцами, что могло приводить к возникновению популяций типа Схул. В этом первичном западном очаге возникали протоевропеоиды и протонегроиды. Протомонголоиды связаны в своем происхождении с западносибирским центром, они Мигрировали в Центральную Азию, где дали начало монголоидной расе; далее они распространялись на восток, где самостоятельно развивалась веддо-австрало-айноидная ветвь неоантропов.
Независимое формирование сапиенса в нескольких (3—4) центрах объясняется кибернетическим механизмом обратной связи между развитием мозга и культуры. Примечательная особенность схемы Тома (в отличие, например, от полицентризма Вейденрейха) в том, что согласно ей эволюция происходит в пределах одного вида—Н. sapiens, к которому он относит не только неоантропов и палеоантропов, но и архантропов (“теория внутривидовой радиации”). Стройность является сильным местом этой схемы, однако следует все же отметить, что по археологическим материалам следы миграции из Передней Азии в Европу в эпоху мустье не установлены.
К числу традиционных полицентристских концепций, предусматривающих происхождение современных больших рас от местных палеоантропов, относится и гипотеза Ф. Смита (1985), согласно которой выделяются 5 центров сапиентации с независимым возникновением Н. Sapiens от местных неандертальцев в Северной Африке, и южнее Сахары, Западной Азии, Восточной Азии, Европе.
В отечественной антропологии превалируют гипотезы “дицентризма” с выделением двух первичных очагов сапиентации: западного (европеоидно-негроидного) и восточного (монголоидного или австрало-монголоидного). В основе полицентристских концепций отечественных антропологов (В.П. Алексеев, А.А. Зубов и др.) и археологов лежит признание мощной центростремительной роли социогенеза на поздних этапах человеческой эволюции. Основные аргументы полицентризма — отдельные краниологические признаки большого таксономического веса, свидетельствующие о хронологической и морфологической преемственности гоминид в ряде локусов Афроевразии, а также археологические материалы, которые не дают достаточных оснований для выделения единого центра возникновения верхнепалеолитической культуры неоантропов. Вообще картина перехода от среднего палеолита к верхнему оказалась довольно сложной. В среднем палеолите существовало множество независимых самобытных культур, различавшихся по происхождению и темпам развития, но отражавших постепенный характер перехода к верхнему палеолиту. Происходило в целом синхронное развитие каменной индустрии среднего палеолита в Африке, Европе и Азии (Ульрих, 1987).
В то же время было бы, конечно, крайностью утверждать, что число локусов сапиентации равно числу очагов перехода от среднего палеолита к верхнему. Жесткой связи “неандерталец—мустье” или “сапиенс—верхний палеолит” не было: первые сапиенсы появились, как уже говорилось, в мустьерскую эпоху, а неандерталец мог преодолевать верхнепалеолитический рубеж, как это доказывает недавняя находка классического неандертальца в Сен-Сезере (Франция) с ранней верхнепалеолитической культурой шатель-перрон. Разумеется, не все эволюционирующие в направлении сапиенса палеоантропы достигли (в силу разных причин) этого уровня. Темпы сапиентации и конечная судьба эволюционирующей популяции еще во многом зависели от конкретной экологической, социодемографической, эпидемиологической ситуации, от особенностей локальных экониш.
Из предшествующего обзора средне- и верхнеплейстоценовых находок гоминид в Афроевразии следует значительный полиморфизм палеоантропов, который на первый взгляд может использоваться как аргумент в пользу моноцентризма, поскольку изменчивость сапиенса признается более слабой. Следует, впрочем, заметить, что при подобных сравнениях индивидуальная изменчивость формирующихся гоминид обычно сопоставляется с групповой вариабельностью современного человека, которая заведомо ниже. К тому же и полиморфизм ранних неоантропов был достаточно велик. Так, в пределах только европейского континента в верхнем палеолите выделяются до 7 “рас”. Наиболее распространенным вариантом был кроманьонский, возникший около 35000—30000 лет назад. Иногда, впрочем, всех ранних неоантропов верхнего палеолита называют “кроманьонцами”. Однако наряду с собственно кроманьонским типом, получившим это наименование по гроту Кро-Маньон во Франции (1868 г.), и близкими к нему гоминидами из гротов Ментоны (Южная Франция), возможно, Младеч (бывшая Чехоcловакия) и других мест, выделяют и отличные варианты, как Комб-Каппель (Франция), “негроидный” (Гримальди, Франция, или Костенки, бывшая СССР), группу центрально- и восточноевропейских неоантропов (Брно— Пшедмости), тип Шанселад (Франция) и др.
Разумеется, расами их можно называть лишь условно, так как эти находки не обнаруживают связи с определенным ареалом и современными антропологическими типами Европы. Можно думать, что таксономическая ценность разграничительных расовых признаков в верхнем палеолите отличалась от современной.
В. В. Бунак развивал точку зрения о краниологическом (и остеологическом) полиморфизме верхнепалеолитических людей, представлявших смесь вариантов полиморфного вида, еще не распавшегося на расы.
Серия черепов из верхнего палеолита и мезолита Европы, обработанная методом главных компонент, не обнаруживает скоплений сходных вариантов и представляется довольно аморфной (Хенке, 1983).
Моноцентристы, опирающиеся преимущественно на материалы по мономерным маркёрам (митохондриальная ДНК), признают сравнительно позднее происхождение рас как локальных вариантов единой популяции, широко расселившейся впоследствии по всем континентам, за счет миграций, изоляции и генного дрейфа.
В пределах полицентристских концепций возможны разные варианты взаимоотношений процессов сапиентации и расогенеза: параллелизм обоих процессов, большая древность рас по сравнению с сапиенсом или, напротив, большая древность процесса сапиентации. Соответственно наряду с традиционным “глубоким” полицентризмом—концепцией о независимом возникновении больших рас Н. sapiens от локальных групп архантропов или палеоантропов в Африке, Европе и Азии и их изолированном развитии в течение длительного времени (до 1 млн лет)—в последнее время все большее распространение получают гипотезы “умеренного” полицентризма, признающие происхождение рас от единого вида (или подвида) Н.sapiens.
В этом случае принципиальные различия моно- и полицентризма относительно эволюционного уровня выделения больших рас в значительной степени нивелируются, так как речь уже идет о поздней внутривидовой дифференциации единого вида или даже подвида на более или менее обширных территориях. При этом обращает на себя внимание чрезвычайно широкое расселение верхнепалеолитических неоантропов, которые обнаруживаются около 40 000—30 000 лет назад в регионах, весьма удаленных друг от друга (Западная Европа, Северная и Южная Африка, Юго-Восточная и Восточная Азия и др.).
Ныне становится все более очевидным, что формирование основных современных рас человечества — европеоидной, негроидной и монголоидной — совсем необязательно связывать с “собственными” архантропами и даже палеоантропами, как это предусматривалось традиционной полицентристской гипотезой. Современные расы скорее всего возникли в ходе внутривидовой дифференциации уже сложившегося раннего сапиенса, тогда как ископаемые гоминиды предшествовавших эпох антропогенеза внесли сравнительно небольшой вклад в специфические расовые комплексы. В нижнем и среднем палеолите появлялись лишь единичные расовые признаки (например, лопатообразность боковых резцов у монголоидов, выступающий нос у европеоидов и др.), и даже в верхнем палеолите еще не было консолидированных современных расовых комплексов, а только их зачатки. Сами же большие расы современного человечества, видимо, возникли лишь в постпалеолитическое время. Это означает, что сапиенс древнее больших рас, процессы сапиентации и расогенеза совпадали лишь частично.
Так, в расовом отношении верхнепалеолитическое население Европы было еще недостаточно дифференцированным. Европеоидный тип установился с неолита, но некоторые его основные черты, как уже говорилось, прослеживаются ранее. Достоверных свидетельств присутствия монголоидов в Восточной Азии в донеолитическую эпоху фактически нет, хотя в Северной Азии они, возможно, обитали уже в верхнем палеолите, как показывает фрагмент черепа из стоянки Афонтова Гора на Енисее и статуэтки из Мальты и Бурети. В Америку человек проник не позже 30000 лет назад, а может быть, и ранее (до 70000 лет), причем в этот период здесь также еще не было сложившегося монголоидного типа. Австралийский континент и Большая Австралия, очевидно, тоже заселялись “недеференцированными” неоантропами в период от 40 000 до 20 000 лет; древнейшая известная из этого региона находка (Лейк- Мунго) имеет возраст около 26 000 лет Позднеплейстоценовые ископаемые черепа из Юго-Восточной Азии (Вьетнам) занимают в морфологическом отношении промежуточное положение между австаралоидами и монголоидами.
Гипотезы как моно-, так и полицентризма основываются на общепризнанном в антропологии положении моногенизма, т. е. концепции, согласно которой все современные расы человечества представляют собой подразделения единого вида. Н.sapiens.
Список литературы
Для подготовки данной работы были использованы материалы с сайта http://flogiston.ru/
Похожие работы
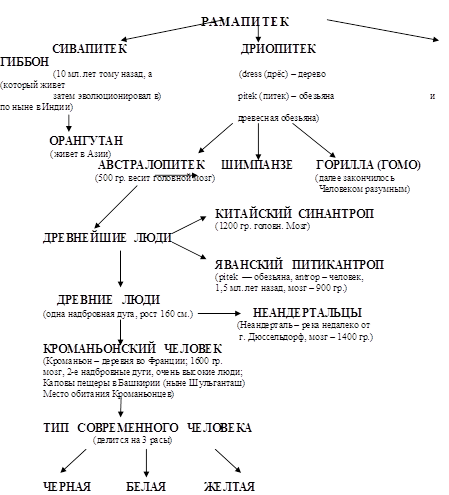

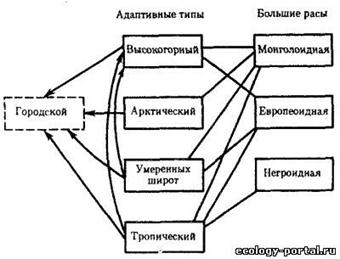

... важнейших вопросов антропологии, этнографии, демографии, исторической и медицинской географии. Расы - это территориальные группы людей, выделяемые на основе их генетического родства, которое проявляется в определенном физическом сходстве. Большинство отечественных антропологов различают три большие расы человечества - монголоидную ("желтую"), европеоидную ("белую") и негроидно-австролоидную (" ...
0 комментариев