Навигация
Возникновение биосинтеза белка
69904
знака
2
таблицы
16
изображений
2.2. Возникновение биосинтеза белка
Далее на основе мира РНК должно было происходить становление механизмов биосинтеза белка, появление разнообразных белков с наследуемой структурой и свойствами, компартментализация систем биосинтеза белка и белковых наборов, возможно, в форме коацерватов и эволюция последних в клеточные структуры - живые клетки.
Проблема перехода от древнего мира РНК к современному белок-синтезирующему миру - наиболее трудная даже для чисто теоретического решения. Возможность абиогенного синтеза полипептидов и белковоподобных веществ не помогает в решении проблемы, так как не просматривается никакого конкретного пути, как этот синтез мог бы быть сопряжен с РНК и подпасть под генетический контроль. Генетически контролируемый синтез полипептидов и белков должен был развиваться независимо от первичного абиогенного синтеза, своим путем, на базе уже существовавшего мира РНК. В литературе предложено несколько гипотез происхождения современного механизма биосинтеза белка в мире РНК, но, пожалуй, ни одна из них не может рассматриваться как детально продуманная и безупречная с точки зрения физико-химических возможностей. Представленная ниже версия наиболее точно отражает процесса эволюции и специализации РНК, ведущего к возникновению аппарата биосинтеза белка (рис. 3), но и она не претендует на законченность.
Предлагаемая гипотетическая схема содержит два существенных момента, кажущихся принципиальными.
1. Постулируется, что абиогенно синтезируемые олигорибонуклеотиды активно рекомбинировали посредством механизма спонтанной неэнзиматической трансэстерификации , приводя к образованию удлиненных цепей РНК и давая начало их многообразию. Именно этим путем в популяции олигонуклеотидов и полинуклеотидов и могли появиться как каталитически активные виды РНК (рибозимы), так и другие виды РНК со специализированными функциями (см. рис. 3). Более того, неэнзиматическая рекомбинация олигонуклеотидов, комплементарно связывающихся с полинуклеотидной матрицей, могла обеспечить сшивание (сплайсинг) фрагментов, комплементарных этой матрице, в единую цепь. Именно таким способом, а не катализируемой полимеризацией мононуклеотидов, могло осуществляться первичные копирование (размножение) РНК. Разумеется, если появлялись рибозимы, обладавшие полимеразной активностью, то эффективность (точность, скорость и продуктивность) копирования на комплементарной матрице должна была значительно возрастать.
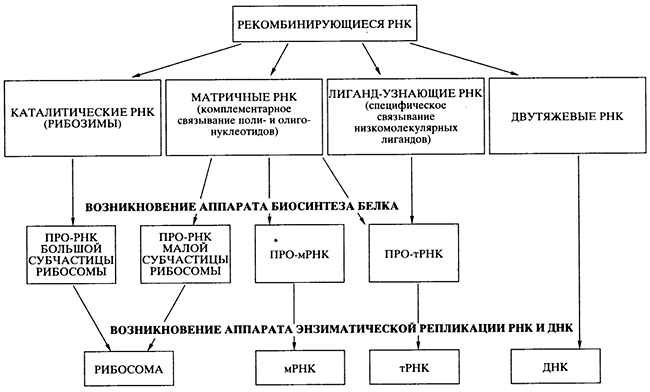
Рис. 3. Схема эволюции и специализации молекул РНК
в процессе перехода от древнего мира РНК к современному миру
генетически детерминированного биосинтеза белков
2. Первичный аппарат биосинтеза белка возник на базе нескольких видов специализированных РНК до появления аппарата энзиматической (полимеразной) репликации генетического материала - РНК и ДНК. Этот первичный аппарат включал:
ü каталитически активную прорибосомную РНК, обладавшую пептидил-трансферазной активностью;
ü набор про-тРНК, специфически связывающих аминокислоты или короткие пептиды;
ü другую прорибосомную РНК, способную взаимодействовать одновременно с каталитической прорибосомной РНК, про-мРНК и про-тРНК (см. рис. 3).
Такая система уже могла синтезировать полипептидные цепи. Среди прочих каталитически активных белков - первичных ферментов (энзимов) - появились и белки, катализирующие полимеризацию нуклеотидов - репликазы, или НК-полимеразы.
Впрочем, возможно, что гипотеза о древнем мире РНК как предшественнике современного живого мира так и не сможет получить достаточного обоснования для преодоления основной трудности - научно правдоподобного описания механизма перехода от РНК и ее репликации к биосинтезу белка. Имеется привлекательная и детально продуманная альтернативная гипотеза А.Д. Альтштейна (Институт биологии гена РАН), в которой постулируется, что репликация генетического материала и его трансляция - синтез белка - возникали и эволюционировали одновременно и сопряжено, начиная с взаимодействия абиогенно синтезирующихся олигонуклеотидов и аминоацил-нуклеотидилатов - смешанных ангидридов аминокислот и нуклеотидов. Но это уже следующая сказка... ("И Шахразаду застигло утро, и она прекратила дозволенные речи".)
3. НУКЛЕИНОВЫЕ КИСЛОТЫ
3.1. Состав нуклеиновых кислот
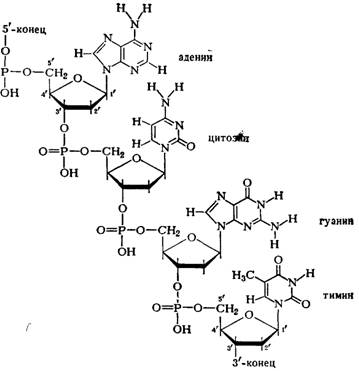
Нуклеиновые кислоты - это биополимеры, макромолекулы которых состоят из многократно повторяющихся звеньев - нуклеотидов. Поэтому их называют также полинуклеотидами. Важнейшей характеристикой нуклеиновых кислот является их нуклеотидный состав. В состав нуклеотида - структурного звена нуклеиновых кислот - входят три составные части:
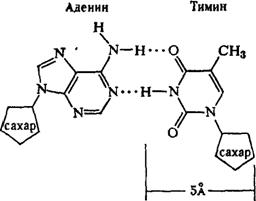
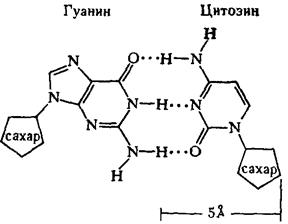
· азотистое основание - пиримидиновое или пуриновое. В нуклеиновых кислотах содержатся основания 4-х разных видов: два из них относятся к классу пуринов и два – к классу пиримидинов. Азот, содержащийся в кольцах, придает молекулам основные свойства.
· моносахарид - рибоза или 2-дезоксирибоза. Сахар, входящий в состав нуклеотида, содержит пять углеродных атомов, т.е. представляет собой пентозу. В зависимости от вида пентозы, присутствующей в нуклеотиде, различают два вида нуклеиновых кислот – рибонуклеиновые кислоты (РНК), которые содержат рибозу, и дезоксирибонуклеиновые кислоты (ДНК), содержащие дизоксирибозу.
· остаток фосфорной кислоты. Нуклеиновые кислоты являются кислотами потому, что в их молекулах содержится фосфорная кислота.
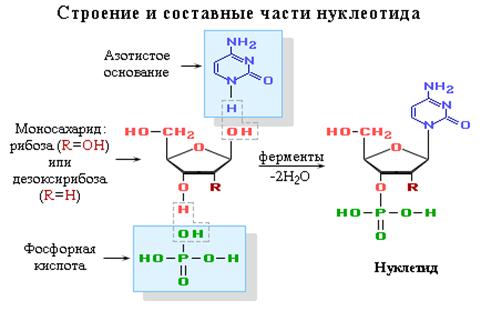
Нуклеотид - фосфорный эфир нуклеозида. В состав нуклеозида входят два компонента: моносахарид (рибоза или дезоксирибоза) и азотистое основание.
В конце 40-х — начале 50-х годов, когда появились такие методы исследования, как хроматография на бумаге и УФ-спектроскопия, были проведены многочисленные исследования нуклеотидного состава НК (Чаргафф, А. Н. Белозерский). Полученные данные позволили решительно отбросить старые представления о нуклеиновых кислотах, как о полимерах, содержащих повторяющиеся тетрануклеотидные последовательности (так называемая тетрануклеотидная теория строения ПК, господствовавшая в 30—40-е годы), и подготовили почву для создания современных представлений не только о первичной структуре ДНК и РНК, но и об их макромолекулярной структуре и функциях.
Метод определения состава ПК основан на анализе гидролизатов, образующихся при их ферментативном или химическом расщеплении. Обычно используются три способа химического расщепления НК. Кислотный гидролиз в жестких условиях (70%-ная хлорная кислота, 100°С, 1ч или 100%-ная муравьиная кислота, 175 °C, 2 ч), применяемый для анализа как ДНК, так и РНК, приводит к разрыву всех N-гликозидных связей и образованию смеси пуриновых и пиримидиновых оснований. При исследовании РНК могут использоваться как мягкий кислотный гидролиз (1 н. соляная кислота, 1OO°C, 1 ч), в результате которого образуются пуриновые основания и пирамидиповые нуклеозид-2'(3')-фосфаты, так и щелочной гидролиз (0,3 н. едкий кали, 37 °С, 20 ч), дающий смесь нуклеозид -2' (3') -фосфатов.
Поскольку в НК число нуклеотидов каждого вида равно числу соответствующих оснований, для установления нуклеотидного состава данной НК достаточно определить количественное соотношение оснований. Для этой цели из гидролизатов с помощью хроматографии на бумаге или электрофореза (когда в результате гидролиза получают нуклеотиды) выделяют индивидуальные соединения. Каждое основание независимо от того, связано оно с углеводным фрагментом или нет, обладает характерным максимумом поглощения в УФ, интенсивность которого зависит от концентрации. По этой причине, исходя из УФ-спектров выделенных соединений, можно определить количественное соотношение оснований, а следовательно, и нуклеотидный состав исходной НК.
При количественном определении минорных нуклеотидов, особенно таких неустойчивых, как дигидроуридиловая кислота, пользуются ферментативными методами гидролиза (ФДЭ змеиного яда и селезенки).
Использование описанных выше аналитических приемов показало, что ПК различного происхождения состоят за редким исключением из четырех основных нуклеотидов и что содержание минорных нуклеотидов может меняться в значительных пределах.
Как будет показано далее, при изучении нуклеотидного состава ДНК были получены данные, которые помогли установить ее пространственную структуру.
Похожие работы
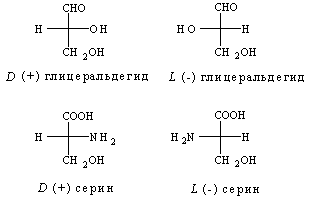
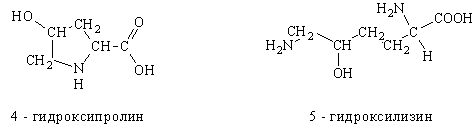
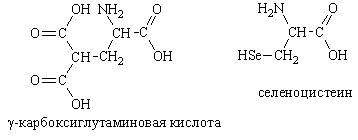
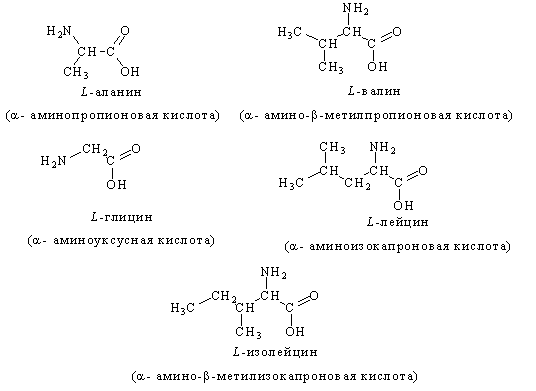
... частью содержится в цитоплазме клеток. Содержание РНК, как правило, в 5-10 раз больше, чем ДНК. Соотношение РНК/ДНК в клетках тем выше, чем интенсивнее в них синтез белка. Нуклеиновые кислоты обладают сильно выраженными кислотными свойствами и при физиологических значениях рН несут высокий отрицательный заряд. В связи с этим в клетках организмов они легко взаимодействуют с различными катионами и ...
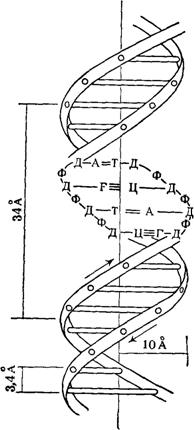
... остатки, соединяющиеся водородными связями с комплементарными основаниями матричной цепи. Рост синтезирующейся цепи происходит в направлении 3'-ОН — 3'-ОН-концам, антипараллельно матричной цепи. Репликация ДНК приводит к удвоению количества генетического материала клетки и, как правило,— к клеточному делению. Поэтому репликация происходит тем чаще, чем короче время генерации вируса или бактерии и ...
... с агрессивным поведением иммунной системы. 21 Медицинская биотехнология, ее задачи и достижения. Биотехнология представляет собой область знаний, которая возникла и оформилась на стыке микробиологии, молекулярной биологии, генетической инженерии, химической технологии и ряда других наук. Рождение биотехнологии обусловлено потребностями общества в новых, более дешевых продуктах для ...
... . ЛИТЕРАТУРА: 1. Курс низших растений //Под ред. М. В. Горленко – М.: Высш. школа, 1981. 2. Жизнь растений, т. 1–3, - М.: Просвещение, 1974-77. 3. Н. Грин, У. Стаут, Д. Тейлор. Биология, т. 1-3. – М.: Мир, 1990. 4. Н.П.Горбунова , Е.С.Клюшникова, Н.А.Комарницкий. и др. Малый практикум по низшим растениям. – М.: Высш. школа, 1976. 5. Русанов В. А. Низшие растения. Методические указания ...
0 комментариев