КРАТКАЯ
ХАРАКТЕРИСТИКА
ФИТОГОРМОНОВ
Механизм
действия цитокининов
Причины и последствия
влияния засоления
на растительные
организмы
Механизмы
адаптации к
засолению
Растения
засолённых
почв – галофиты
Методы
исследований
Влияние
6-БАП на растения
кукурузы в
зависимости
от уровня засоления
Величина
отрицательного
влияния засоления
находится в
прямой зависимости
от концентрации
соли в субстрате
Навигация
КРАТКАЯ ХАРАКТЕРИСТИКА ФИТОГОРМОНОВ
Влияние 6-БАП на растения кукурузы при разном уровне засоления
90994
знака
10
таблиц
19
изображений
1. КРАТКАЯ ХАРАКТЕРИСТИКА ФИТОГОРМОНОВ.
Важнейшими представителями эндогенных регуляторов роста растений являются фитогормоны. Это вещества, которые синтезируются в растениях, транспортируются по ним в малых концентрациях и способны вызывать ростовые или формативные эффекты 10.
Все фитогормоны обладают тремя основными особенностями:
1. Эндогенное происхождение. Изменения в интенсивности синтеза того или иного фитогормона, вызванное внутренними или внешними причинами, вызывает ответную реакцию растения — переход к другому характеру ростовых или формативных процессов.
2. Возможность транспортировки по растению. Физиологический смысл этой особенности состоите том, что фитогормон, образовавшийся в одном органе (например, в апикальной меристеме стебля), должен обладать свойством регуляции ростовых процессов в других органах (например, в корне). Именно таким образом достигается взаимодействие органов и целостность растения
3. Способность в малых концентрациях (10-12 - 10-7М) вызывать заметные ростовые или формативные эффекты. Примером ростового эффекта может служить ускорение или замедление роста стебля, формативного — дефолиация.
Работы многих ученых 3, 7, 9, 21, 30, 33 показали, что фитогормоны участвуют в регуляции обмена веществ на всех этапах жизни растений — от развития зародыша до полного завершения жизненного цикла и отмирания. Они определяют характер роста и развития растений, формирования новых органов, габитуса, цветения, старения вегетативных частей, перехода к покою и выхода из него и т.п.
В настоящее время известно восемь групп фитогормонов: пять из которых относятся к классическим группам - ауксины, гиббереллины, цитокинины, абсцизовая кислота, этилен и три открытые сравнительно недавно - брассиностероиды, жасминовая и салициловая кислоты. Все они могут активизировать, но могут и тормозить функциональную активность клеток. Общим условием для действия любого фитогормона является наличие в клетках специфичных рецепторов. Все фитогормоны вызывают у компетентных клеток сравнительно быстрые физиологические реакции, связанные, очевидно, с мембранами и более медленные изменения, зависящие от синтеза белков и нуклеиновых кислот 33.
Предполагается наличие в растениях еще неизвестных классов гормонов, о свойствах которых можно судить только по косвенным показателям. Это — факторы цветения (флориген, ВЕНВ), ризогенеза и роста клеток (фузикокцин), повышения устойчивости растений (олигосахариды). Все это свидетельствует о том, что гормональная система растений значительно сложнее, чем представлялось до недавнего времени.
Помимо естественных фитогормонов, получено большое количество синтетических аналогов этих природных соединений, которые часто обладают высокой физиологической активностью. Строго говоря, эти вещества не могут быть отнесены к фитогормонам, так как не образуются в растениях, однако многие из них по активности не уступают фитогормонам или даже превосходят их 32.
2. ОБЩАЯ ХАРАКТЕРИСТИКА ЦИТОКИНИНОВ.
2.1. Химическая структура.
Цитокининами называется один из типов фитогормонов, обладающих определенной совокупностью биологической активности, которая весьма разнообразна и проявляется при регуляции роста, органообразования, процессов старения и покоя.
Цитокинины принимают участие наряду с другими фитогормонами в регуляции самых разнообразных физиологических процессов в растении. Для них, как и для других фитогормонов, характерна полифункциональность. Кроме того, действие цитокининов появляется в неразрывной связи с действием других фитогормонов и природных ингибиторов.
В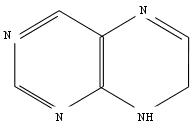 химическом
отношении
природные
цитокинины
и их синтетические заменители представляют собой производные
6-аминопурина
с заместителем
в аминогруппе
при шестом
атоме углерода
пуринового
кольца.
химическом
отношении
природные
цитокинины
и их синтетические заменители представляют собой производные
6-аминопурина
с заместителем
в аминогруппе
при шестом
атоме углерода
пуринового
кольца.
Пурин
Основными представителями цитокининового ряда являются: кинетин, 6-бензиламинопурин (6-БАП), 8-азакинетин, бензимидазол, которые принадлежат к синтетическим цитокининам, а представителем природных цитокининов является зеатин, который был выделен из растения кукурузы 32, 48.
Цитокинины пуринового ряда слаборастворимы в воде, но хорошо растворимы в этаноле, этиловом эфире, ряде щелочей и кислот. Цитокинины устойчивы к нагреванию, автоклавированию, действию щелочей и кислот. Образование природных цитокининов происходит в корневой системе, а передвижение в надземные органы идёт по ксилеме 49.
2.2. История открытия.
Цитокинины были открыты в 1955 г. Скугом и Миллером с сотрудниками в Висконсинском университете в США. Их обнаружению помог так называемый “дефектный объект”, которым явился каллюс сердцевины стебля табака. Он образовался на кусочках сердцевины стебля в условиях стерильной культуры, но быстро прекращал рост в связи с исчерпанием какого-то фактора, исходно присутствовавшего в тканях стебля. Ввиду того, что вернуть активный рост каллюса удавалось добавлением к питательной смеси, содержащей ИУК, дрожжевого экстракта, а так же кокосового молока, Скугом и сотрудниками была предпринята попытка выделить недостающие для роста каллюса вещество из дрожжевого экстракта. При этом выяснилось, что активное вещество обладает свойствами пурина.
Вместе с тем, проверка природных пуриновых оснований, так же как гидрометода РНК и ДНК показала, что они не способны обеспечить рост стеблевого каллюса табака. Неожиданно активным оказался эфирный экстракт из препарата ДНК сельди. Впоследствии выяснилось, что активное вещество образуется в любом препарате ДНК при ее деградации путем автоклавирования в кислой среде. Это вещество было выделено в виде кристаллов и идентифицировано химически. Оно оказалось 6- фурфуриламинопурином, который был назван кинетином.
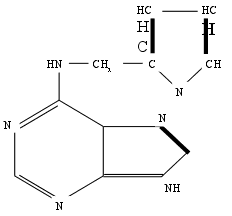
6-фурфуриламинопурин
Принцип химического синтеза производных пурина с замещением в аминогруппе у шестого атома углерода пуринового кольца был известен ранее, поэтому вслед за химической идентификацией кинетина последовал его синтез. Затем было синтезировано много других активных соединений, которые отличались от кинетина характером заместителя в аминогруппе при шестом атоме углерода. Все эти соединения, включая кинетин, были объединены под общим названием кинины. Это название было распространено в литературе до 1965 г. Однако ввиду того, что в физиологии животных термин “кинины” еще ранее был применен для совершенно иных соединений - биологически активных полипептидов, Скуг, Стронг и Миллер предложили заменить название кинины на цитокинины. В настоящее время термин цитокинины стал общепризнанным. 22.
2.3.Физиологическая роль цитокининов.
Для цитокининов, как и для других фитогормонов характерна многофункциональность. Также необходимо отметить, что в большинстве случаев физиологическое действие цитокининов проявляется в непосредственной связи с действием других фитогормонов.
а) Стимуляция деления клеток.
Стимуляция клеточного деления представляет собой одно из характерных свойств цитокининов, она была показана на различных растительных объектах: изолированных тканях, кончиках корня, растущих листьях, семядолях, точках роста стебля, зародыша прорастающих семян, однако, некоторые растительные объекты могут быть не компетентны для реакции на цитокинин активацией клеточных делений 22.
Особенно ярко проявляются процессы деления на культуре изолированных тканей. Так, в работах Скуга с сотрудниками было показано, что у изолированной сердцевинной ткани стебля табака в стерильной культуре одна ИУК несколько усиливала синтез ДНК, вызывала в отдельных клетках митозы, но не индуцировала клеточных делений. Точно так же один кинетин не вызывал деления клеток. В его присутствии не происходили митозы, а синтез ДНК он стимулировал в меньшей степени, чем ИУК. Только совместное действие ауксинов и кинетина значительно активировало синтез ДНК, вызывало митозы и индуцировало деление, клеток в изолированной сердцевине стебля табака.
В последующем были предприняты попытки разграничить во времени действие ИУК и кинетина в индукции клеточных делений у этой ткани. Полученные данные не имеют пока однозначной интерпретации, но позволяют предполагать, что начальные стадии процесса индуцируются одним ауксином, тогда как в последующем необходимы оба гормона.
В интактном растении наблюдается корреляция между активность цитокининов и скоростью клеточного деления. Предполагают, что цитокинины стимулируют цитокинез - последнюю стадию деления клетки.
Достаточное эндогенное содержание цитокининов, нехватка ауксинов или других необходимых веществ могут стать причиной отсутствия стимулирующего действия цитокининов на деление клеток. Кроме того, в случае высокого эндогенного содержания цитокининов их предоставление извне может привести и к угнетению клеточных делений 24.
б) Влияние цитокининов на рост клеток.
Уже в первых работах по цитокининам на стеблевых каллюсах табака в лаборатории Скуга было показано, что цитокинины влияют не только на деление клеток, но и на их рост растяжением. Скуг пришел к выводу о том, что для роста клеток так же, как и для их деления, нужны и цитокинины, и ауксины, причем концентрации этих веществ и соотношение между ними, стимулирующее рост клеток, отличается от тех, которые стимулируют клеточное деление.
Наиболее активно цитокинины стимулируют увеличение размера клеток в зонах растущих листьев двудольных травянистых растений и их изолированных семядолей. Важно отметить, что с помощью цитокининов также можно стимулировать рост в тканях, уже давно закончивших этот процесс. На отрезках листьев однодольных растений стимуляция роста клеток цитокинином не удается. Помимо листьев, стимуляция роста клеток цитокининами показана и у многих других объектов.
Однако отмечены случаи, когда цитокинины угнетали рост клеток. Угнетение чаще всего обнаруживается на отрезках стебля и корнях, причем цитокинины снимают стимулирующее действие на рост стебля ауксинов. Не исключено, что различие роста клеток листьев и клеток стебля и корня при воздействии цитокининов определяется различием в диапазоне стимулирующих концентраций.
Цитокинины активируют в высечках синтез необходимых для роста РНК. Они составляют небольшую часть от общего содержания РНК в клетке. В их состав должны входить иРНК, кодирующие необходимые для процесса роста белки. Трансляция этих РНК происходит в цитоплазме на 80 S рибосомах, в результате чего увеличивается новообразование структурных и ферментных белков, в том числе белков лимитирующих ростовые процессы. Вследствие этого фитогормон активирует в зависимости от типа ткани деление и рост клеток, или процессы, связанные с дифференцированием 21. Однако после того как синтезируется некоторый запас необходимых для роста белков, стимуляция роста цитокинином становится независимой от дальнейшего образования белка до тех пор, пока лимитирующие рост белки не будут использованы в клетках 49.
в) Действие цитокининов на органогенез.
Еще одним важным свойством цитокининов является участие в процессах органогенеза у растений. Индукция с помощью цитокининов органогенеза у недифференцированной ткани стеблевого каллюса табака была впервые показана Скугом с сотрудниками, которые с помощью ИУК и кинетина вызывали образование у каллюса корней и побегов и установили, что для закладки каждого из этих органов требовались свои специфические концентрации обоих фитогормонов. Для дифференциации корней требовалось присутствие 2 мг/л ИУК и 0,02 мг/л кинетина. Повышение концентрации кинетина до 0,5 - 1 мг/л приводило к индукции формирования стеблевых почек. Таким образом, сдвиг соотношения концентрации ауксин - цитокинин в сторону цитокинина способствует образованию стеблевых почек, а в сторону ауксина - закладке корней 10.
Меняя содержание фитогормонов в питательной среде, можно было направлять органогенез в сторону образования корней или побегов. При этом в ряде случаев цитокинины, стимулируя образование побегов, вместе с тем задерживали закладку корней. Так как концентрации цитокининов, стимулирующие закладку корней, крайне низки, в большинстве объектов хватает, по-видимому, собственных цитокининов для осуществления этого процесса. Возможно, именно этим объясняется тот факт, что данные извне цитокинины ингибируют корнеобразование 22.
Цитокинины положительно влияют также на закладку боковых корней у изолированных участков корня, на черешках изолированных листьев фасоли и др. 32.
Существенно отметить, что с помощью цитокининов удается также влиять на закладку и дифференциацию генеративных органов, вызывая зацветание растений в условиях неблагоприятного температурного или фотопериодического режима, а также повлиять на пол цветков.
Цитокинины участвуют, также в регуляции органогенеза у споровых растений - папоротников и мхов. Под действием цитокининов у протонемы мха, представляющей собою нитевидное образование, индуцируется закладка стеблевых почек.
Таким образом, цитокинины оказывают большое влияние на процесс дифференциации, характер которого меняется в зависимости от концентрации цитокинина, его соотношения с ауксином и особенностей объекта 37.
г) Прерывание покоя и стимуляция прорастания семян под действием цитокининов.
Еще одно свойство цитокининов - это способность прерывать покой спящих почек древесных растений, клубней (бегония), семян некоторых видов, в частности семян древесных пород. Отмечено, что в ряде случаев обработка экзогенными цитокининами оказывает на семена, почки и клубнелуковицы действие, подобное обработке холодом. При пониженных температурах во время стратификации семян в них происходит значительное увеличение содержания цитокининов, что способствует их прорастанию; поэтому обработка семян экзогенными цитокинином имитирует действие холода. Кратковременное замачивание в растворе 6-бензиламинопурина низкой концентрации повышает всхожесть семян, ослабленную в результате их длительного хранения.
Также стимулирующее действие цитокининов на прорастание обнаружено на семенах латука, которые нуждаются для индукции этого процесса в действии красного света. Цитокинины стимулируют их прорастание, как в темноте, так и при кратковременном воздействии красным светом 32.
д) Влияние цитокининов на рост целых растений.
Цитокинины оказывают влияние не только на рост клеток, но и на рост всего растения в целом. В литературе имеются данные, что под влиянием опрыскивания синтетического аналога цитокинина кинетином сухой вес проростков ячменя и пшеницы возрастает на 8-12% 33. В опытах с проростками кукурузы было показано, что опрыскивание кинетином увеличивает объем корневой системы, сырой и сухой вес надземных органов 34. В условиях полевого опыта на растениях озимой пшеницы опрыскивание синтетического аналога цитокинина 6-БАП увеличило темпы роста и продуктивность, повышало число зерен в колосе 35.
Кинетин, добавленный в крайне низких концентрациях к раствору Кнопа, стимулировал рост проростков подсолнечника, фасоли и люпина, растений ряски в темноте, а также проростков редиса на свету. Погружение растений табака на 1 час корнями в раствор цитокинина (5х10-4 М) вызывало значительное усиление роста листьев на растении и повышало в них содержание калия. Есть указания о возможности повлиять с помощью цитокининов на рост плодов яблони и на формирование урожая у растений кукурузы 19, 29, 44, 45.
е) Защитное действие цитокининов при неблагоприятных факторах среды.
Цитокинины повышают устойчивость клеток к самым различным неблагоприятным воздействиям, таким как, действие повышенной и пониженной температуры, обезвоживание, грибная и вирусная инфекция, механическое воздействие и влияние различных химических агентов 7.
Причины защитного действия цитокининов в каждом конкретном случае могут быть различными и, очевидно, требуют специального изучения. Однако не исключено, что механизм такого действия может оказаться и одинаковым. Например, он может проявляться через действие цитокининов на структурное и функциональное состояние различных макромолекулярных компонентов клетки и, в частности, на состояние их мембранного аппарата. Электронно-микроскопические исследования показывают, что цитокинины задерживают деградацию различных мембранных структур в клетках листа, включая ламеллы стромы и граны хлоропластов, мембранные структуры митохондрий, эндоплазматический ретикулум. При помощи цитокинина удавалось предотвратить разрушение в срезанных листьях мембранной оболочки сферосом, содержащих гидролитические ферменты, и таким путем защитить от разрушения белки, нуклеиновые кислоты и липоиды плазмы. По-видимому, действие цитокининов на мембранный аппарат клетки может иметь существенное значение в осуществлении их регуляторного действия на обмен веществ растений, и не исключено, что имеет отношение к защитному действию цитокининов при различных неблагоприятных воздействиях.
Иную интерпретацию может иметь защитное действие цитокининов на листья целых растений в условиях повышенной температуры и засухи. По-видимому, в таких случаях в листьях может возникать дефицит эндогенных цитокининов за счет снижения их поступления из корней, что и создает условия для защитного действия данных извне цитокининов. Правда, необходимо упомянуть, что такое защитное действие проявляется не всегда.
В работе Кабузенко и Горшенкова показано, что влияние хлоридного засоления на активность корневой системы пшеницы и кукурузы оказывает отрицательное воздействие. А обработка 6-БАП на фоне засоления субстрата положительно влияет на метатическую активность меристемы кончиков корней проростков кукурузы и пшеницы. Было установлено, что под воздействием 6-БАП нивелировалось действие соли, приближая продолжительность фаз клеточного цикла к уровню контроля. Внесение 6-БАП в среду проращивания способствовало сокращению продолжительности клеточного цикла меристемы корня в целом на 25% по сравнению с растениями, прорастающими при «чистом» засолении. Под влиянием цитокинина произошло существенное уменьшение продолжительности интерфазы (на 28%) и профазы (на 29,1%) митоза. Последующие фазы (метафаза, анафаза и телофаза), связанные с формированием и функцией веретена деления, при действии 6-БАП, проходили медленнее, чем на фоне «чистого засоления». Таким образом, наличие цитокинина в среде может способствовать снятию ингибирующего влияния засоляющих ионов на прохождение интерфазы и первой фазы митоза в меристеме корней злаков 13.
В литературе имеются данные о действии водного стресса, засоления, а также сверхоптимальных температур на прорастание семян. В результате чего в семенах происходит снижение эндогенных цитокининов с одновременным повышением уровня АБК. Обработка экзогенным цитокинином, по-видимому, способствует восстановлению нормального фитогормонального баланса в клетке.
Полученные Калининой и сотрудниками морфометрические данные свидетельствуют о том, что засоление снижает как сырой, так и сухой вес растений кукурузы, а также длину наземной и подземной части проростка. Длина наземной части растений при засолении была ниже контрольных на 55%, корня – на 45%, сырая масса растений меньше на 50%, сухая – на 60%. Наличие в среде проращивания 6-БАП способствует нормализации ростовых процессов. В варианте NaCl+6-БАП длина стебля увеличилась на 40% по сравнению с «чистым» засолением, корня – на 25%; сырая масса проростков возросла на 20%, а сухая – на 32% 14.
Данные, полученные Калининой и Кабузенко, свидетельствуют о значительном снижении содержания белка в корнях трёхдневных проростков кукурузы на солевом фоне: содержание белков понизилось на 44%. Добавление в солевую среду регуляторов роста способствовало увеличению содержания белка в корнях проростков. Позитивное действие наиболее чётко было выражено при добавлении в среду проращивания растений кукурузы препарата 6-БАП, который увеличивал содержания белка в корнях на 30% как на бессолевом фоне, так и в условиях засоления.
Пероксидазе отводится важная роль в процессе утилизации накапливающихся при стрессе метаболитов, в частности Н2О2 2. В результате проведенных опытов было установлено, что наличие хлорида натрия в среде проращивания увеличивало активность пероксидазы в корнях проростков в 2,3 раза по сравнению с контролем. Изменение активности пероксидазы в корнях кукурузы в условиях хлоридного засоления можно считать проявлением нарушений нормальных метаболических процессов в клетках, которое может быть снивелировано действием экзогенных цитокининов. Применение 6-БАП в условиях засоления способствовало снижению активности этого фермента на 69% 15.
Таким образом, можно сказать, что цитокинины оказывают на растения положительное влияние при любых неблагоприятных условиях среды, таких как: хлоридное засоление, действие световых и температурных воздействий, водного стресса, повышенной засухи.
Похожие работы

... по фосфору, азоту или сахару. Режим хемостата позволяет с помощью фиксированной скорости разбавления поддерживать константную скорость деления и плотность клеток в популяции. Клеточные суспензии в биотехнологии используются для получения вторичных метаболитов, многие из которых являются ценными лекарственными препаратами, для промышленного выращивания клеточной биомассы и для клеточной селекции. ...
0 комментариев