Навигация
Введение
Что такое сон, для чего он нужен организму? Вопрос о функциональном назначении столь обыденного состояния кажется настолько наивным, что даже не требует раздумий: конечно, для отдыха! Однако такой ответ порождает цепочку новых вопросов: что такое отдых? Почему он столь продолжителен и столь сложно организован? Почему приурочен к определенным периодам суток? Почему для отдыха недостаточно телесного покоя, а необходимо еще и выключение органов чувств, что, казалось бы, резко повышает уязвимость по отношению к неблагоприятным факторам среды? Почему теплокровные животные, у которых «постоянство внутренней среды является залогом свободной жизни», вынуждены, подобно своим холоднокровным предкам, каждые сутки на несколько часов впадать в состояние неподвижности и ареактивности?
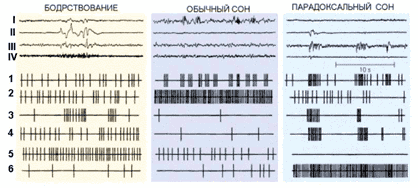
На протяжении многих столетий сон рассматривался именно по этим внешним признакам, то есть состоянию покоя и пониженной реактивности. Такому подходу не смогло помешать даже формирование представлений о двух состояниях «внутри» естественного сна, принципиально отличных друг от друга и от бодрствования (медленно волновая и парадоксальная фазы). Однако в последнее время появляется все большее число фактов, которые не укладываются в такие представления. Так, в начале 80-х годов сотрудники I Московского медицинского института В.С. Ротенберг и С.И. Кобрин, изучая сон больных с полной атрофией мышечной системы, не выявили его сокращения, хотя эти больные вовсе не нуждались в соматическом (телесном) «отдыхе». Значит, сон не есть покой, а телесный отдых вовсе не обязательный элемент физиологического сна.
Аналогичным образом можно рассмотреть и такую общепринятую характеристику сна, как ареактивность, то есть психическую заторможенность, отсутствие реакции на внешние стимулы. Во-первых, это «апостериорный» признак сна, поскольку порог пробуждения можно определить, лишь разбудив человека. Во-вторых, ареактивность, так же как и неподвижность, не служит достаточным признаком, поскольку она характерна для целого ряда заболеваний и других патологических состояний: фармакологического сна, наркоза, комы и прочего.
1. Природа сна
1.1 Теории сна
Гуморальная теория, в качестве причины сна рассматривает вещества, появляющиеся в крови при длительном бодрствовании. Доказательством этой теории служит эксперимент, при котором бодрствующей собаке переливали кровь животного, лишенного сна в течение суток. Животное-реципиент немедленно засыпало. В настоящее время удалось идентифицировать некоторые гипногенные вещества, например пептид, вызывающий дельта-сон. Но гуморальные факторы не могут рассматриваться как абсолютная причина возникновения сна. Об этом свидетельствуют наблюдения за поведением двух пар неразделившихся близнецов. У них разделение нервной системы произошло полностью, а системы кровообращения имели множество анастомозов. Эти близнецы могли спать в разное время: одна девочка, например, могла спать, а другая бодрствовала.
Ретикулярная теория сна и бодрствования. В ретикулярной формации ствола мозга находится множество нейронов, аксоны которых идут почти ко всем областям головного мозга (кроме неокортекса). В конце 1940-х годах Моруцци и Мэгуном было обнаружено, что высокочастотное раздражение ретикулярной формации ствола мозга кошек приводит к их мгновенному пробуждению. Повреждение ретикулярной формации вызывает постоянный сон, перерезка же сенсорных трактов такого эффекта не дает. Ретикулярную формацию стали рассматривать как область головного мозга, участвующую в поддержании сна. Сон наступает, когда ее активность пассивно, либо под действием внешних факторов падает. Активация ретикулярной формации зависит от количества сенсорных импульсов, поступающих в нее, а так же от активности нисходящих волокон между передним мозгом и стволовыми структурами. Однако позднее было установлено, что: ретикулярная формация вызывает не только бодрствования, но и сон, что зависит от места наложения электродов при стимуляции ее электрическим раздражителем; нейронное состояние ретикулярной формации в бодрствующем состоянии и во время сна мало, чем отличается; ретикулярная формация является не единственным центром бодрствования: они так же представлены и в медиальном таламусе, и в переднем гипоталамусе.
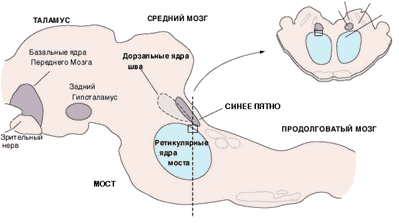
Серотонинергическая теория сна и бодрствования. В верхних отделах ствола мозга обнаружены две области: ядро шва и голубое пятно. Медиатором в клетках ядра шатра является серотонин, а голубого места – норадреналин. В конце 1960-х Жуве пришел к выводу, что эти две нейронные системы принимают в участии возникновении сна. Разрушение ядер шва у кошки приводит к полной бессоннице в течение нескольких дней, за несколько последующих недель сон восстанавливается. Частичная бессонница может быть вызвана подавлением синтеза серотонина хлорфенилаланином, введением предшественника серотонина ее можно устранить. Разрушение голубого пятна приводит к полному исчезновению БДГ-сна, но не влияет на медленный сон. Истощение запасов серотонина вызывает бессонницу, а введение предшественников серотонина нормализует только медленный сон. Все это позволило предположить, что серотонин приводит к торможению структур, ответственных за бодрствование. Было установлено, что голубое пятно подавляет импульсацию ядра шва, и это ведет к пробуждению.
Сейчас доказано, что нейроны ядер шва выделяют серотонин во время бодрствования: он служит медиатором в процессе пробуждения и «гормоном сна» в бодрствующем состоянии: стимулируя выход вещества сна, который вызывает сон. Фаза БДГ-сна обеспечивается подголубоватым ядром. Было показано, что сон и бодрствование определяются активацией специфических центров головного мозга. Одни из таких центров является ретикулярная формация, которая расположена в стволе мозга. Он из основных компонентов ретикулярной формации являются холинергические ядра, расположенные на уровне мосто-среднемозгового сочленения. Нейроны этих ядер имеют высокий уровень активности во время бодрствования и REM-фазы и инактивированны во время медленного сна.
В регуляции процессов сна-бодрствования принимают участие и другие эргические системы головного мозга, медиаторами которых являются: серотонин, норадреналин, гистамин, глутамат, вазопрессин. Вероятно, что диссомнии обусловлены нарушением функционирования нейротрансмиттерных систем.
Подкорковая и корковая теории сна: при различных опухолевых или инфекционных поражениях подкорковых, особенно стволовых, образований мозга, у больных отмечаются различные нарушения сна – от бессонницы до длительного летаргического сна, что указывает на наличие подкорковых центров сна. При раздражении задних структур субталамуса и гипоталамуса животные засыпали, а после прекращения раздражения они просыпались, что указывает на наличие в этих структурах центров сна. В лаборатории И.П. Павлова было установлено, что при длительной выработке тонкого дифференцировочного торможения животные часто засыпали. Поэтому ученый рассматривал сон как следствие процессов внутреннего торможения, как углубленное, разлитое, распространившееся на оба полушария и ближайшую подкорку торможение (корковая теория сна).
Однако ряд фактов не могли объяснить ни корковая, ни подкорковая теории сна. Наблюдения за больными, у которых отсутствовали почти все виды чувствительности, показали, что такие больные впадают в состояние сна как только прерывается поток информации от действующих органов чувств. Например, у одного больного из всех органов чувств был сохранен только один глаз, закрытие которого погружало больного в состояние сна. Многие вопросы организации процессов сна получили объяснение с открытием восходящих активирующих влияний ретикулярной формации ствола мозга на кору больших полушарий. Экспериментально было доказано, что сон возникает во всех случаях устранения восходящих активирующих влияний ретикулярной формации на кору мозга. Были установлены нисходящие влияния коры мозга на подкорковые образования. В бодрствующем состоянии при наличии восходящих активирующих влияний ретикулярной формации на кору мозга нейроны лобной коры тормозят активность нейронов центра сна заднего гипоталамуса. В состоянии сна, когда снижаются восходящие активирующие влияния ретикулярной формации на кору мозга, тормозные влияния лобной коры на гипоталамические центры сна снижаются.
Между лимбико-гипоталамическими и ретикулярными структурами мозга имеются реципрокные отношения. При возбуждении лимбико-гипоталамических структур мозга наблюдается торможение структур ретикулярной формации ствола мозга и наоборот. При бодрствовании за счет потоков афферентации от органов чувств активируются структуры ретикулярной формации, которые оказывают восходящее активирующее влияние на кору больших полушарий. При этом нейроны лобных отделов коры оказывают нисходящие тормозные влияния на центры сна заднего гипоталамуса, что устраняет блокирующие влияния гипоталамических центров сна на ретикулярную формацию среднего мозга. При уменьшении потока сенсорной информации снижаются восходящие активирующие влияния ретикулярной формации на кору мозга. В результате чего устраняются тормозные влияния лобной коры на нейроны центра сна заднего гипоталамуса, которые начинают еще активнее тормозить ретикулярную формацию ствола мозга. В условиях блокады всех восходящих активирующих влияний подкорковых образований на кору мозга наблюдается медленно волновая стадия сна.
Гипоталамические центры за счет связей с лимбическими структурами мозга могут оказывать восходящие активирующие влияния на кору мозга при отсутствии влияний ретикулярной формации ствола мозга. Эти механизмы составляют корково-подкорковую теорию сна (П.К. Анохин), которая позволила объяснить все виды сна и его расстройства. Она исходит из того, что состояние сна связано с важнейшим механизмом – снижением восходящих активирующих влияний ретикулярной формации на кору мозга. Сон бескорковых животных и новорожденных детей объясняется слабой выраженностью нисходящих влияний лобной коры на гипоталамические центры сна, которые при этих условиях находятся в активном состоянии и оказывают тормозное действие на нейроны ретикулярной формации ствола мозга.
Сон новорожденного периодически прерывается только возбуждением центра голода, расположенного в латеральных ядрах гипоталамуса, который тормозит активность центра сна. При этом создаются условия для поступления восходящих активирующих влияний ретикулярной формации в кору. Эта теория объясняет многие расстройства сна. Бессонница, например, часто возникает как следствие перевозбуждения коры под влиянием курения, напряженной творческой работы перед сном. При этом усиливаются нисходящие тормозные влияния нейронов лобной коры на гипоталамические центры сна и подавляется механизм их блокирующего действия на ретикулярную формацию ствола мозга. Длительный сон может наблюдаться при раздражении центров заднего гипоталамуса сосудистым или опухолевым патологическим процессом. Возбужденные клетки центра сна непрерывно оказывают блокирующее влияние на нейроны ретикулярной формации ствола мозга.
Иногда во время сна наблюдается так называемое частичное бодрствование, которое объясняется наличием определенных каналов реверберации возбуждений между подкорковыми структурами и корой больших полушарий во время сна на фоне снижения восходящих активирующих влияний ретикулярной формации на кору мозга. Например, кормящая мать может крепко спать и не реагировать на сильные звуки, но она быстро просыпается даже при небольшом шевелении ребенка. В случае патологических изменений в том или ином органе усиленная импульсация от него может определять характер сновидений и быть своего рода предвестником заболевания, субъективные признаки которого еще не воспринимаются в состоянии бодрствования.
Диафферентационная теория сна и бодрствования. В конце 1930-х годов Бремер обнаружил, что ЭЭГ кошки с перерезкой, отделяющей спинной мозг от головного после восстановления от операционного шока демонстрирует циклические чередования, характерный для сна-бодрствования. Если перерезка произведена на уровне четверохолмия, то есть исключены сенсорные стимулы, кроме зрительных и обонятельных, наблюдается типичная для сна ЭЭГ. Бремер пришел к выводу, что ЦНС индуцируется и поддерживается: для бодрствования необходимо минимума сенсорной стимуляции, сон – состояние, характеризующееся, прежде всего, снижением эффективности сенсорной стимуляции мозга, что подтверждает теорию пассивного бодрствования.
Однако: во-первых – в изолированном переднем мозге со временем проявляются ритмические колебания, характерные для ритма сон-бодрствования. Кроме того, изоляция человека в звуконепроницаемой камере приводит к уменьшению длительности сна. Во-вторых – данные о влиянии коры на состояние бодрствования неверны, так как циркадные ритмы сна-бодрствования наблюдаются и у новорожденных детей аэнцефаликов.
Эндогенная теория сна. Человек ощущает определенную потребность во сне, что связывают с наличием факторов сна, циркулирующих в крови. Тогда во время сна должны восстанавливаться их нормальные концентрации. Предполагается, что факторы сна накапливаются во время бодрствования до вызывающего сон уровня. Согласно другой гипотезе эти факторы накапливаются во время сна: образуются и выделяются. Из мочи и спиномозговой жидкости был выделен гликопептид – дельта-пептид, вызывающий медленный сон при введении другим животным. Есть фактор и БДГ-сна. Вторая гипотеза привела к открытию в крови пептида дельта-сна, вызывающего глубокий сон.
Однако найденные факторы вызывают сон у человека и только у некоторых видов животных. Кроме того, он может возникнуть и под действием других видов веществ. На сегодняшний день неизвестно, какую физиологическую роль в процессе выполняют найденные факторы.
Фармакологический сон неадекватен по своим механизмам естественному сну. Снотворные препараты ограничивают активность разных структур мозга – ретикулярной формации, гипоталамической области, коры головного мозга. Это приводит к нарушению естественных механизмов формирования стадий сна, нарушению процесса консолидации памяти, переработки и усвоения информации.
Похожие работы
... клеток коры головного мозга. Таким образом, постоянная связь человека с внешним миром это главное условие его нормальной сознательной деятельности, которая является конечным результатом всех материальных процессов, происходящих в головном мозге. ПАМЯТЬ, ЕЕ ЗНАЧЕНИЕ И ФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ Память — способность живых существ воспринимать, отбирать, хранить и использовать информацию для ...

... , Манассеина пришла к выводу, что сон для организма важнее пищи. В 1889 году М. Манассеина опубликовала большую книгу под названием: «Сон как треть жизни, или физиология, патология, гигиена и психология сна». Эта книга явилась настоящей энциклопедией, где впервые в популярном изложении приводились все знания того времени по проблеме сна. Книга имела огромный успех, была переведена на основные ...
... информации. Отбор информации осуществляется благодаря реакции внима- ния,рассмотренной ранее.Остановимся теперь более подробно на механизмах формирования энграммы. Важный вклад в создание современной теории памяти внес канадский физиолог Хебб.Он предположил,что внешний стимул сразу приводит к образованию лабильного следа памяти в моз- ге,который вскоре исчезает.Переход информации в долговремен- ...
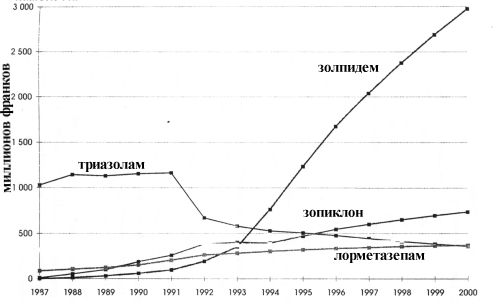
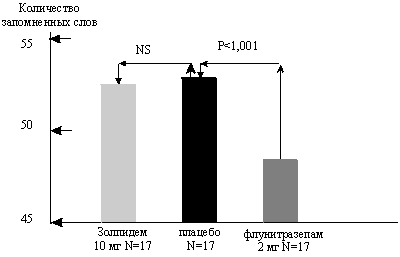

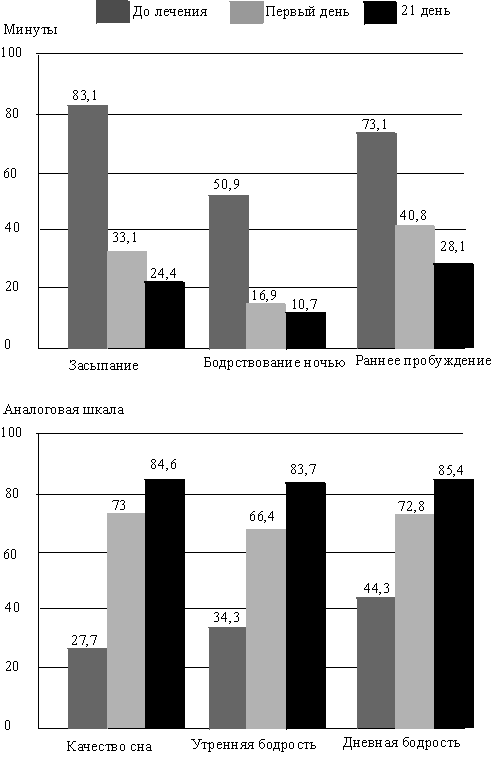
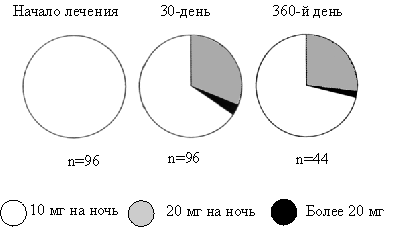
... даже на нее достоверного влияния, чего нельзя сказать о производном бензодиазепина ( рис. 2). Рис.5 Доля пациентов, получавших различные дозы золпидема при лечении нарушений сна в течение года Утреннее пробуждение после приема ивадала носит характер свежести после полноценного сна. Помимо короткого периода полувыведения препарата, помимо его селективности в отношении омега-1 рецепторов, это ...
0 комментариев