Навигация
3.3 Природній відбір
В процесі видоутворення природний відбір переводить випадкову індивідуальну мінливість в біологічно корисну групову – популяційну, видову. Стабілізуюча його форма викликає «вдалі» комбінації алелей від попередніх етапів революції. Відбір підтримує також стан генетичного поліморфізму. Зміна біологічних факторів історичного розвитку соціальними призвела до того, що в людських популяціях відбір втратив функцію відображення. За ним збереглись функції стабілізації генофонду і підтримання спадкового різноманіття.
На користь дій стабілізуючої форми природного відбору говорить, наприклад, велика смертність серед недоношених і переношених новонароджених в порівнянні з доношеними. Напрямок відбору серед таких дітей залежить від зниження загальної життєздатності. Негативний відбір за одним лукусом можна проілюструвати на прикладі системи груп крові «резус» (Rh).
Близько 85% населення Європи має в еритроцитах антиген Rh і утворює групу Rh-позитивних індивідуумів. Решта людей із європейських популяцій не мають цього антигену і являються Rh-негативними. Синтез антигену Rh контролюється домінуючою алеллю D і відбувається в осіб з генотипами DD і Dd. Резус-негативні люди являються рецесивними гомозиготами (dd). При вагітності Rh-негативної жінки (dd) Rh-позитивним плодом (чоловік DD чи Dd, плід Dd) при порушенні цілісності плаценти при родах Rh-позитивні еритроцити плоду проникають в організм матері і імунізують його. При наступній вагітності Rh-позитивним плодом (Dd) анти-Rh-антитіла проникають через плаценту в організм і розрушують його еритроцити. Розвивається гемолітична хвороба новонародженого. Основним її симптомом являється тяжка анемія.
В наш час медицина в змозі боротися з цією формою патології у вигляді негайного переливання новонародженому Rh-негативної крові чи введення анти-Rh-антитіл для запобігання імунізації матері. В випадку відсутності медичної допомоги новонароджений нерідко помирав. При Rh-негативному фенотипі матері Rh-позитивний плід завжди гетерозиготний (Dd). Це означає, що зі смертністю такого індивідууму із генофонду популяції, до якого він належить, вилучається рівна кількість домінантних і рецесивних алелей лукусу «резус». Відбір в даному випадку направлений проти гетерозигот.
Під дією відбору знаходяться алелі, що контролюють синтез антигенів системи груп крові АВ0. про це свідчить зниження проти очікуваної кількості дітей в АВ0-несумісних шлюбах жінок 0-групи з чоловіками А, В або АВ-груп [Додаток Є].
Особливість відбору залу кусами групоспецифічних антигенів еритроцитів, таких, як «резус» або АВ0, полягає в тому, що він діє на рівні зиготи чи ранній стадії ембріогенезу. Помічено, що в шлюбах, де батько і мати несумісні за фактором «резус», кількість дітей з гемолітичною хворобою менша очікуваної. Так, серед білого населення США Rh-негативні люди складають приблизно16%. В такій популяції частота алеля d рівна 40%, а вірогідність розвитку гемолітичної хвороби за випадкового підбору шлюбних пар – 9%. Насправді частота цієї хвороби складає 1:150 – 1:200. Одним із факторів зниження являється одночасна несумісність батьків за лукусом АВ0. В цьому випадку Rh-позитивні еритроцити плоду, що потрапили в організм матері, руйнуються і імунізація не відбувається.
Негативний відбір діє в більшості популяцій людей за алелями аномальних гемоглобінів. Його особлива жорстокість обумовлюється тим, що він направлений проти гомозигот. Дитина, що помирає, наприклад, від серповидно-кліткової анемії, являється гомозиготною за алеллю S. Кожна така смерть видаляє із генофонду популяції алелей одного виду. Це призводить до досить швидкого зниження мінливості за відпорним лукусом. В багатьох популяціях людей частота алелей аномальних гемоглобінів, втому числі і S, не перевищує 1%.
Висока частота алелей таких аномальних гемоглобінів, як S, C, D, E, в деяких регіонах планети ілюструє дію природного відбору за підтриманням в людських популяціях стану балансованого генетичного поліморфізму. Негативний відбір по відношенню до алеля S перекривається могутнім позитивним відбором гетерозигот HbAHbS завдяки високій життєдіяльності останніх в ядрах тропічної малярії.
Дослідження в Уганді показали, що кількість збудників в 1 мл крові залежить від генотипу дитини і складає до 10 000 у HbSHbS, до 160 00 у HbAHbS і до 800 000 у HbAHbA. В північній Греції дослідили 48 сімей, в яких спостерігались і серповидно-клітковість і малярія. Серед братів і сестер, хворих серповидно-клітковою анемією, хворіло малярією 16 із 25 з генотипами HbAHbA і 1 із 23 з генотипами HbAHbS.
Таким чином, нормальні люди хворіли в 13 раз частіше, ніж індивідууми з аномальним генотипом. Знешкодження фактору контрвідбору призводить до зниження частоти алеля серповидно-клітковості. Цією причиною, що діє уже протягом декількох століть поряд з метисацією, пояснюють відносно низьку частоту гетерозигот HbAHbS серед північноамериканських негрів (8-9%) в порівнянні з африканськими (близько 20%).
В наведених прикладах діям негативного відбору, що знижує в генофондах деяких популяцій людей концентрацію певних алелей, протистоять контрвідбори, які підтримують частоту цих алелей на достатньо високому рівні. Результатом накладення численних і різнонаправлених векторів відбору являється формування і підтримання генофондів популяцій в стані, що забезпечує утворення в кожному поколінні генотипів достатньої пристосованості з врахуванням місцевих умов. Завдяки соціально-економісним перетворенням, успіхам лікувальної і особливо профілактичної медицини вплив відбору на генетичний склад популяцій людей прогресивно знижується.
ВИСНОВКИ
На відміну від змін прокаріотичного геному перетворення генома в еволюції еукаріот пов’язані з наростаючим збільшенням кількості ДНК. Це збільшення спостерігається в процесі прогресивної еволюції еукаріот. На фоні такого збільшення більша частина ДНК являється «мовчазливою», тобто не кодує амінокислот в білках чи послідовностях нуклеотидів в рРНК і тРНК. В складі ДНК знаходяться високо і помірно повторювані послідовності. Вся маса ДНК розподілена між певною кількістю спеціалізованих структур – хромосом. Хромосоми на відміну від нуклеотиду прокаріот мають складну хімічну організацію. Еукаріоти в більшості випадків диплоїдні. Час генерації у них значно більший, ніж у прокаріот. Відмічені особливості, що оформилися у ході еволюції геному еукаріот, допускають широкі структурні зміни і забезпечують не лише адаптивну (приспособлювальну), але і прогресивну еволюцію.
Серед перечислених вище моментів збільшення розмірів геному в еволюції еукаріот звертає на себе особливу увагу. Цей процес може здійснюватися різноманітними шляхами. Найрізкіше розмір геному змінюється в результаті поліплоідизації, яка достатньо широко розповсюджена в природі. Її суть полягає у збільшенні кількості ДНК і хромосом, кратних галоїдному. Досягнуте в результаті стану поліплоїдії призводить до збільшення дози всіх генів і складає залишок «сирого» генетичного матеріалу, який нарешті видозмінюється внаслідок мутації і відбору.
В ході революції в результаті накопичення мутацій і дивергенції нуклеотидних послідовностей поліплоїдизація супроводжувалась переходом до диплоїдного стану. Само собою збільшення дози генів ще не означає досягнення однозначно позитивного біологічного результату. Про це свідчить розвиток в еволюції еукаріотів механізмів компенсації дози генів, що збільшується, в ході їх експресій шляхом скорочення тривалості життя в клітинах зрілої РНК. Так, у тетраплоїдних карпових риб у відповідь на збільшення дози генів рРНК в молекулах рРНК соматичних клітин утворюються приховані внутрішні розриви, які призводять до передчасного їх старіння і скорочення складу в цитоплазмі.
Якби збільшення об’єму геному відбувалося тільки в результаті поліплоїдизації, то в природі мала б спостерігатися стрибкоподібна змін його розмірів. Насправді цей процес демонструє плавне збільшення складу ДНК в геномі. Це дозволяє допустити можливість існування інших механізмів, що змінюють його об’єм.
Дійсно, деякі значення у визначенні об’єму геному являються мають хромосомні перебудови, що супроводжуються змінами складу ДНК у них, такі, як дуплікації, делеції і транслокації. Вони обумовлюють повтор, втрату деяких послідовностей в складі хромосоми або перенесення їх в інші хромосоми.
Важливим механізмом збільшення об’єму геному являється ампліфікація нуклеотидних послідовностей, яка полягає в утворення їх копій, що призводить до виникнення повторюваних участків ДНК. Особливістю геному еукаріотів являється наявність таких повторів в великій кількості, що свідчить про суттєвий внесок механізму ампліфікації в збільшення розмірів спадкового матеріалу. Ампліфіціровані послідовності утворюють сімейства, в яких вони зібрані разом (тандемна організація) або ж розподілені по різних хромосомах. Конкретні зміни, що призводять до ампліфікації, бувають різноманітними. Поява тандемів повторюваних послідовностей пояснюється, наприклад, нерівним кросинговером, внаслідок якого виникають багаторазові дуплікації окремих участків ДНК. Можлива ампліфікація шляхом вирізання фрагменту в наступній його реплікації поза хромосомою і влаштуванням копій в інші хромосоми. Допускають також ампліфікацію, що здійснюється шляхом «зворотної транскрипції» ДНК та РНК за участі ферменту зворотної транскриптази з наступним влаштуванням копій ДНК в різні лукуси хромосом. В усіх випадках ампліфікація деякої послідовності призводить до виникнення в геномі більш чи менш численних повторів і полягає некратному збільшенню його об’єму. Наявність таких поворотів в поєднанні з мутаційним процесом являється початковим ступенем дивергентної еволюції однотипних послідовностей в межах сімейства з відповідною зміною якостей кодованих білків чи РНК.
Ярким прикладом еволюційної долі ампліфікованих нуклеотидних послідовностей являються сімейства глобінових генів, що широко розповсюджені в природі у видів різних рівнів організації. У вищих хребетних відомий ряд глобинових генів, що контролюють синтез поліпептидів гемоглобіну. Людина в геномі має вісім активних глобинових генів, що утворюють два сімейства. Сімейство генів, що визначають синтез α-глобінів, що мають ξ-глібинові гени, які експресуються у плода дорослої людини. Це сімейство генів розміщується в 16-й хромосомі в наступному порядку: 5΄—ξ²—ξ¹—α²—α¹—3΄. Сімейство генів, що визначають синтез β-глобінів, що розміщені в 11-й хромосомі, має ε-глобіновий склад ген ембріона, два схожі γ-глобінові гени Gγ і Аγ, малий δ- і великий β-глобінові гени дорослих: 5΄—ε—Gγ—Aγ—δ—β—3΄.
Вивчення гомології продуктів вказаних генів і генотипів міоглобіну у різних видів організмів дозволило допустити спільність походження цих сімейств. Ймовірно, близько 1100 млн. років тому відбулась дуплікація гену-попередника, що дала початок гемоглобіновим і міоглобіновим генам. Пізніше, близько 500 років тому, на початковому етапі революції хребетних відбулась дуплікація, яка давала початок двом (α і β) сімействам глобі нових генів, що супроводжувалася транс локацією. Близько 200 млн. років тому наступна дуплікація призвела до виникнення в сімействі β-глобінових генів плодів і дорослих. Близько 100 млн. років тому відбулось утворення ε- і γ-глобінових генів і , нарешті, 40 млн. років тому зявились і δ- і β-глобінові гени.
Сімейства α- і β-глобінові гени організовані в кластери, що, напевне, виникли в результаті тандемної дуплікації генів. В складі вказаних кластерів поряд з активно функціонуючими на різних стадіях онтогенезу генами виявлені неактивні, чи псевдогени. Останні, напевне, виникли в результаті появи в них змін, несумісних з можливістю їх експресії. В сімействі β-глобінових генів є два псевдогени: ψβ¹ і ψβ². В α-сімействі – один псевдоген.
Дивергенція ампліфікованих послідовностей з утворенням різних генів чи їх сімейств обумовлена накопиченням в них різних змін у вигляді замін генних мутацій. Про гомологію глобі нових генів обох сімейств свідчить наявність у всіх наявних глобі нових генах хребетних двох ін тронних ділянки, що займають в них строго однакове положення. Таку ж організацію мають і псевдо гени ψα¹ у людини, ψα² у кролика. Однак у ψα³-псевдогені миші в ході еволюції обидва інтригони виявились точно вирізаними.
Результатом ампліфікації невеликих послідовностей ДНК в межах функціональної одиниці являється подовження гена, за якого із простих генів можуть утворюватися більш складні. Це може відбуватися за рахунок тандем них дуплікацій. Наприклад, в генах, що кодують варіабельні ділянки імуноглобулінів миші, послідовності із 600 п. н. утворюються в результаті 12 тандемних повторів вихідної послідовності предків в 48 п. н. другим прикладом подовження гену тандем них дуплікацій слугує ген колагена α², який у куриці складається із 34 000 п. н. і має більше 50 екзонів. Довжина таких ділянок у всіх випадках кратна девяти нуклеотидним парам. Еволюція цих екзотів, напевне, ішла від гіпотетичного вихідного блоку довжиною в 54 пари нуклеотидів.
Список використаних джерел
1. Гончаренко М.С., Бойчук Ю.Д. Екологія людини. – К., 2005. – 393 с.
2. Загайко А.Л., Дербильова А.Г., Павільченко Ю.В. Біологія. Довідник абітурієнта. – Харків, 2006. – 286 с.
3. Рудий Б.А. Криза еволюцыонызму. – К., 2005. – 44-51 с.
Яблоков А.В. Популяционная біологія.-М.-Высшая школа-1987.- 142-143с.
4. Яригін. Біологія. Т1, Т2
ДОДАТОК А
Найбільш населені країни в 1998 р. з прогнозом на 2050 р.
| Місце | Країна | Населення, млн чол. 1998 рік | Країна | Населення, млн чол. 2050 рік |
| 1 | Китай | 1255 | Індія | 1 533 |
| 2 | Індія | 976 | Китай | 1 517 |
| 3 | США | 274 | Пакистан | 357 |
| 4 | Індонезія | 207 | США | 348 |
| 5 | Бразилія | 165 | Нігерія | 339 |
| 6 | Росія | 148 | Індонезія | 318 |
| 7 | Пакистан | 147 | Бразилія | 143 |
| 8 | Японія | 126 | Бангладеш | 218 |
| 9 | Бангладеш | 124 | Ефіопія | 213 |
| 10 | Нігерія | 122 | Іран | 170 |
| 11 | Мексика | 96 | Конго | 165 |
| 12 | Німеччина | 82 | Мексика | 154 |
| 13 | В'єтнам | 78 | Філіппіни | 131 |
| 14 | Іран | 73 | В'єтнам | 130 |
| 15 | Філіппіни | 72 | Єгипет | 115 |
| 16 | Єгипет | 66 | Росія | 114 |
| 17 | Туреччина | 64 | Японія | 110 |
| 18 | Таїланд | 62 | Туреччина | 98 |
| 19 | Франція | 60 | ПАР | 91 |
| 20 | Ефіопія | 59 | Таїланд | 89 |
ДОДАТОК Б
Основні індикатори рівня життя населення світу
| Країна | ІЛР, 1997 | Індекс очі-куваності тривалості життя | Індекс рі-вня осві-ти насе-лення | Внутрішній валовий продукт | Індекс розвитку людського потенціалу | Рейтинг країни за ІЛР, 1998 |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 |
| Австрія | 0,904 | 0,87 | 0,95 | 0,90 | 0,908 | 16 |
| Австралія | 0,922 | 0,89 | 0,99 | 0,89 | 0,929 | 4 |
| Алжир | 0,665 | 0,73 | 0,63 | 0,63 | 0,683 | 107 |
| Аргентина | 0,827 | 0,80 | 0,91 | 0,77 | 0,837 | 35 |
| Білорусія | 0,763 | 0,72 | 0,93 | 0,65 | 0,781 | 57 |
| Бельгія | 0,923 | 0,87 | 0,99 | 0,91 | 0,925 П ТЛ7 | 7 ~7А "• |
| Бразилія | 0,739 | 0,70 | 0,83 | 0,70 | 0,747 | 74 : |
| Великобританія | 0,918 | 0,87 | 0,99 | 0,89 | 0,918 | 10 |
| Німеччина | 0,906 | 0,87 | 0,95 | 0,89 | 0,911 | 14 |
| Греція | 0,867 | 0,89 | 0,91 | 0,81 | 0,875 | 25 |
| Данія | 0,905 | 0,84 | 0,96 | 0,91 | 0,911 | 15 |
| Єгипет | 0.616 | 0,69 | 0,59 | 0,57 | 0,623 | 119 |
| Ізраїль | 0,883 | 0,88 | 0,90 | 0,87 | 0,883 | 23 |
| Індія | 0,545 | 0,63 | 0,54 | 0,47 | 0,563 | 128 |
| Іспанія | 0,894 | 0,88 | 0,95 | 0,85 | 0,899 | 21 |
| Італія | 0,900 | 0,89 | 0,93 | 0,89 | 0,903 | 19 |
| Канада | 0,932 | 0,90 | 0,99 | 0,90 | 0,935 | 1 |
| Казахстан | 0,740 | 0,71 | 0,91 | 0,60 | 0,754 | 73 |
| Китай | 0,701 | 0,75 | 0,78 | 0,57 | 0,706 | 99 |
| Латвія | 0,744 | 0,72 | 0,90 | 0,61 | 0,771 | 63 |
| Люксембург | 0,902 | 0,86 | 0,89 | 0,96 | 0,908 | 17 |
| Нідерланди | 0,921 | 0,88 | 0,99 | 0,89 | 0,925 | 8 |
| Норвегія | 0,927 | 0,89 | 0,98 | 0,92 | 0,934 | 2 |
| Польща | 0,802 | 0,79 | 0,92 | 0,70 | 0,814 | 44 |
| Росія | 0,747 | 0,69 | 0,92 | 0,63 | 0,771 | 62 |
| Респ. Корея | 0,852 | 0,79 | 0,95 | 0,82 | 0,854 | 11 |
| Румунія | 0,752 | 0,75 | 0,88 | 0,63 | 0,770 | 64 |
| США | 0,927 | 0,86 | 0,97 | 0,95 | 0,929 | 3 |
| Туреччина | 0,728 | 0,71 | 0,76 | 0,69 | 0,723 | 85 |
| Угорщина | 0,725 | 0,76 | 0,91 | 0,71 | 0,817 | 43 |
| Україна | 0,721 | 0,73 | 0,92 | 0,52 | 0,744 | 78 |
| Фінляндія | 0, 913 | 0,86 | 0,99 | 0,89 | 0,917 | 11 |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 |
| Франція | 0,918 | 0,89 | 0,97 | 0,90 | 0,892 | 12 |
| Чехія | 0,833 | 0,81 | 0,91 | 0,78 | 0,843 | 34 |
| Швейцарія | 0,914 | 0,89 | 0,92 | 0,92 | 0,915 | 13 |
| Швеція | 0,923 | 0,89 | 0,99 | 0,88 | 0,926 | 6 |
| ПАР | 0,695 | 0,50 | 0,87 | 0,72 | 0,697 | 103 |
| Японія | 0,924 | 0,92 | 0,94 | 0,92 | 0,924 | 9 |
| Нігерія | - | - | - | - | 0,293 | 173 |
| Сьєра-Леоне | - | - | - | - | 0,252 | 174 |
ДОДАТОК В
Динаміка якості життя в Росії за період переходу до демократичного ладу (1998-2003 рр.)
| Статистичний показник якості життя | 1986-1987 | 1994 | 2003 | % | Людська «ціна»переходу |
| Смертність (на 1000 жителів) | 10,5 | 15,7 | 16,6 | 158 | 7,7 млн померлих понад норму |
| Загальна народжуваність (на 1000 жителів) | 17,1 | 9,5 | 10,4 | 61 | 14 млн втрачених потенційних громадян |
| Самогубства (на 100 000 жителів) | 23 | 42 | 37 | 161 | 280 тис. додаткових самогубств |
| Вбивства (на 100 000 жителів) | 7,6 | 32 | 30 | 395 | 390 тис. додаткових вбивств |
| Розбійні напади і крадіжки (тис.) | 34 | 187 | 214 | 207 | |
| Розлучення (на 1000 шлюбів) | 410 | 620 | 850 | 207 |
ДОДАТОК Г
Кількість ВІЛ-інфікованих і хворих на СНІД дорослих (від15 до 49 років) на кінець 1999 року
| Регіон | Кількість, чол. |
| Африка на південь від Сахари | 23 400 000 |
| Південна і Південно-Східна Азія | 5 400 000 |
| Латинська Америка | 1 200 000 |
| Північна Америка | 890 000 |
| Східна Азія і Тихоокеанський регіон | 530 000 |
| Західна Європа | 520 000 |
| Східна і Центральна Європа | 410 000 |
| Карибський басейн | 350 000 |
| Північна Африка і Близький Схід | 210 000 |
| Австралія і Нова Зеландія | 15 000 |
ДОДАТОК Ґ
Кількість ВІЛ-інфікованих і хворих на СНІД (від 15 до 49 років) в африканських країнах на кінець 1999 року
| Країна | Кількість, % | Країна | Кількість, % |
| Ботсвана | 38,5 | Кенія | 14,0 |
| Свазіленд | 25,2 | ЦАР | 14,0 |
| Зімбабве | 25,0 | Мозамбік | 13,2 |
| Лесото | 23,5 | Джібуті | 11,7 |
| Замбія | 20,0 | Бурунді | 11,3 |
| ПАР | 20,0 | Руанда | 11,2 |
| Намібія | 19,5 | Кот-д’Івуар | 10,7 |
| Малаві | 16,0 | Ефіопія | 10,6 |
ДОДАТОК Д
Динаміка густоти народонаселення Землі
| Період | Епоха | Густота населення, чол./ 100км2 |
| Сучасність | Індустріальна | 1700 |
| 80 років тому | Індустріальна | 1100 |
| 180 років тому | Аграрно-індустріальна | 620 |
| 130 років тому | Аграрно-індустріальна | 490 |
| 330 років тому | Індустріально-аграрна | 370 |
| 2000 років тому | Аграрна | 100 |
| 6000 років тому | Перші міста | 75 |
| 10 тис. років тому | Мезоліт | 4 |
| 25 тис. років тому | Верхній палеоліт | 4 |
| 30 тис. років тому | Середній палеоліт | 1,2 |
| 100 тис. років тому | Нижній палеоліт | 0,4 |
ДОДАТОК Е
Приклади алелей, що мають адаптивне значення
| Алелі і генотипи | Географічна розповсюдженість | Адаптивне значення |
| Групи крові системи АВО, алель В | Повсюдно, частіше в Азії | Відносна стійкість до чуми |
| Алель А | Повсюдно | Відносна стійкість до язви шлунку і дванадцятипалої кишки |
| Трансферріни – білки, що зв’язують залізо, алель Т1 | Висока частота в зоні тропічного поясу | Резистентність до багатьох інфекційних хвороб |
| Кисла фосфатаза еритроцитів, алель Аср | Висока частота у бушменів і негроїдів Центральної Африки | Висока активність ферменту при підвищеній температурі |
| Алель АсР і група крові АВ | Висока частота на о. Нова Гвінея | Стійкість до тропічної малярії |
| Алель АсР і група крові А або алель АсР і група крові АВ | Висока частота у жителів Чукотки і Аляски | Легкість адаптації в умовах холодного клімату |
| Вушна сіра, алель d | Висока частота на Далекому Сході | При генотипі dd низький рівень холестерину і концентрація ліпідів в крові, висока концентрація лізоциму у вушній сірці |
ДОДАТОК Є
Вплив АВ0-несумісності на плодовитість в ембріонах
| Показник | АВ0-сумісні | АВ0-несумісні |
| Кількість сімей, які були оглянуті | 812 | 617 |
| Викидні,% | 10,3 | 15,3 |
| Бездітні шлюби,% | 9,8 | 18,2 |
| Середня кількість дітей на одну матір | 2,6 | 2,2 |
Похожие работы
... ’язків між ендокринними органами, характерні для старіння зміни імунної системи, процеси старіння сполучної тканини та ін. Вперше термін був запропонований І. І. Мечниковим у 1903 р. Тема 4.: Особливості дії елементарних факторів добору в популяціях людей на сучасному етапі План 1. Вплив на людські популяції елементарних еволюційних факторів. 2. Значення міграцій в екології людини. Характер ...
... Екологічна структура популяції — це її стан на даний момент (кількість та густота особин, їх розміщення у просторі, співвідношення груп за статтю і віком, морфологічні, поведінкові й інші особливості) Структура популяції являє собою форми адаптації до умов її існування, є своєрідним віддзеркаленням природних сил, які на неї впливають. Нинішня структура тієї чи іншої популяції відбиває водночас ...
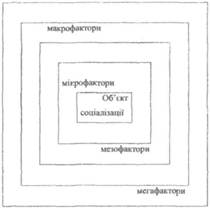
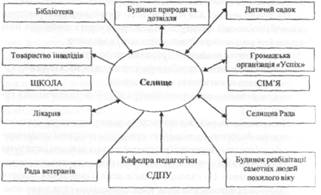
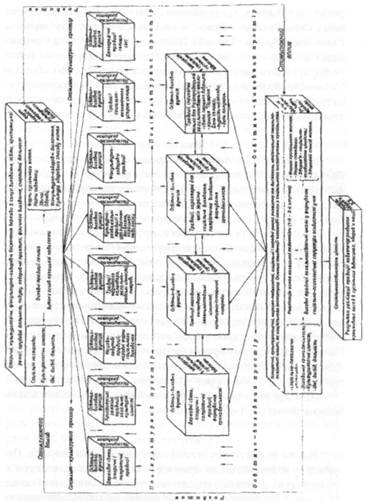
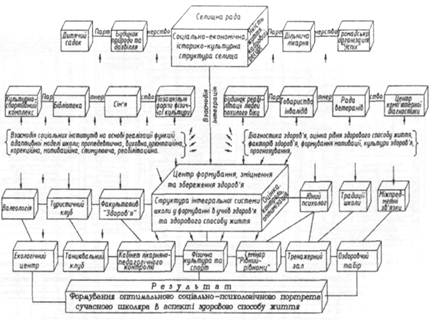

... ів є актуальною, оскільки на її основі реально можна розробити формувальні, розвивальні та оздоровчі структурні компоненти технологічних моделей у цілісній системі взаємодії соціальних інститутів суспільства у формуванні здорового способу життя дітей та підлітків. На основі інформації, яка отримана в результаті діагностики, реалізується методика розробки ефективних критеріїв оцінки інноваційних ...
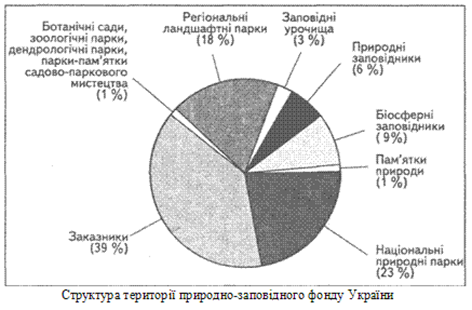
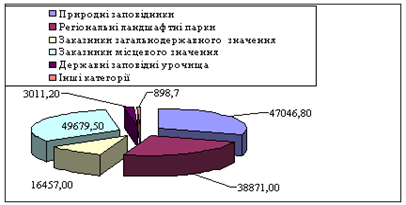
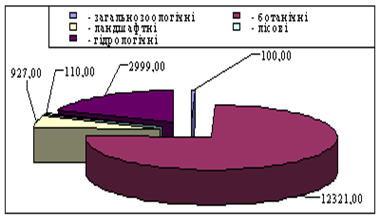
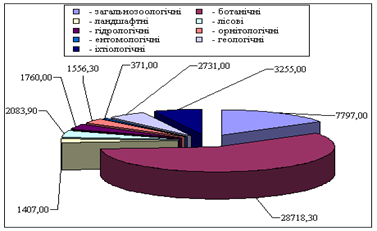
... ідних територій багатогранна і важлива. Вони знаходяться на обліку і під державним контролем, який покладений в Україні на Міністерство по охороні навколишнього середовища. Розділ 2. Заказники та їх роль у збереження біорозмаїття 2.1 Заказники місцевого та державного рівня у структурі природо-заповідних територій Рівненщини Природно-заповідний фонд (ПЗФ) Рівненської області станом ...
0 комментариев