Навигация
2.2.3 ПАРАПОДИАЛЬНЫЙ КОМПЛЕКС
Все сегменты метастомиума, иначе туловищные сегменты, снабжены боковыми выростами — параподиями, вооруженными многочисленными щетинками. Пара-подии в основном служат локомоторными органами, но одновременно в не-которых случаях выполняют и дыхательные функции, а также являются орудиями защиты. Более подробно остановимся на морфологии параподий Aphroditiformia, у которых, как и у большинства эррантных полихет, параподий моностикного характера, т. е. невроподии и нотоподии объединены общим подиальным стволом (в противоположность многим седентарным полихетам, у которых имеются дистикные параподий, характеризующиеся разобщенными ветвями).

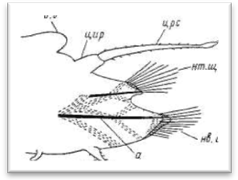
Рис. 8. Усиковая (А) и чешуйчатая (Б) параподии полиноид.
Параподий чешуйчатых червей имеют большей частью сильно вытянутую форму и четко отделены от главного ствола тела самого червя (на брюшной стороне они ограничиваются нефридиальными папиллами). Длина параподий нередко равна ширине тела и даже может ее превосходить. Различают два основных типа параподий: 1) усиковые параподии, снабженные удлиненным верете-образной формы спинным усиком, выполняющим осязательную функцию, и 2) чешуйчатые параподии, у которых спинной усик заменен плоской чешуйкой, или элитрой, выполняющей преимущественно защитную функцию (рис. 8). Параподии снабжены двумя крупными внутренними осевыми щетинками, как правило, темного цвета. Это так называемые ацикулы (aciculum), которые соответствуют двум ветвям параподий: верхней, или спинной (нотоподия— notopodium), и нижней, или брюшной (невроподия — neuropidium). Такие параподии обозначаются как двуветвистые (biramus, biremis), однако верхняя ветвь нередко остается недоразвитой и выражена лишь небольшим бугорком на передне-верхней стороне подиального ствола. Параподии с недоразвитой нотоподией иногда называются «ложнодвуветвистые» или «полуразветвленные» (sub-biramus, sesquiramus). При беглом взгляде последние могут быть приняты за одноветвистые; они наблюдаются преимущественно лишь у многосегментных полиноид типа Arctonoe, Lepidasihenia и Perolepis. Короткие полиноиды — Harmotkoe и большинство других родов имеют, напротив, мощную нотоподию, снабженную большим количеством крупных щетинок, образующих плотный пучок. В качестве «типичных» для чешуйчатых червей принято считать параподии Harmotkoe. Однако за исходный тип параподий для всех полихет мы принимаем одноветвистые. В таксономической литературе при характеристике многих родов полиноид пишется, что параподии у них с редуцированной нотоподией. Нам представляется, что в большинстве случаев мы имеем дело с недоразвитой нотоподией. Последнее, в частности, подтверждается данными по развитию: в онтогенезе нотоподия закладывается позже невроподии.
Невроподия представляет собой непосредственное продолжение главного подиального ствола (truncus), служит как бы базой для нотоподии и несет основную локомоторную функцию. Нижняя (невроподиальная) ацикула доходит до дистального конца подиального выроста, так называемого ацикулярного бугорка (acicular lobe), придавая всей конструкции параподии упругий харак тер. Невроподии на дистальном конце обычно имеют две мягкие, разного размера и формы кожистые лопасти (lamella): переднюю, или предщетинковую (presetal), и заднюю, или послещетинковую (postsetal). Щетинки на невроподии расположены полукругом, причем когда невроподия сильно вытянута и прижата к субстрату, то и большинство ее щетинок также упирается в последний (это обеспечивается разной длиной самих щетинок и степенью их выдвижения). Выдвижение и втягивание щетинок происходит под воздействием специальной мускульной системы — протракторов и ретракторов. Мышечные волокна ретракторов плотно окружают основание ацикул, а таковые протракторов прикреплены к внутренней стенке параподии.
Нотоподия чешуйчатых червей развита различно: у комменсальных полиноид она обычно недоразвита, а у свободно ползающих по субстрату, так же как и у зарывающихся в грунт форм, напротив, нотоподия снабжена мощным пучком щетинок, образующим под элитрами как бы фильтр, предохраняющий респираторный канал на спинной стороне от засорения посторонними детритными и минеральными частицами. Дорсальная ветвь параподии не выполняет какую-либо локомоторную функцию, она обычно представлена в виде небольшой широкой подушечки с многочисленными рядами нотоподиальных щетинок, которые, как правило, сидят не так глубоко, как невроподиалъные щетинки, и к тому же они не втягиваются и не выдвигаются; поддерживающая их дорсальная ацикула значительно тоньше и короче вентральной.
Форма параподии в разных семействах весьма разнообразна (рис. 9), но во всех случаях она хорошо адаптирована к способам передвижения: ползанию-по твердому субстрату («маршированию»), зарыванию в мягкий грунт и обитанию-в трубках или иных убежищах. Параподии Aphrodita (рис. 9,1) вооружены мощными дорсальными щетинками в виде гладких или зазубренных игл, торчащих в разные стороны, и очень длинными волосовидными щетинками, образующими поверх спинных чешуек густой войлок; такое вооружение параподии, с одной стороны, способствует быстрому закапыванию червя передним своим концом в грунт путем поочередных полувинтовых движений тела, а с другой стороны — надежно предохраняет спинную сторону червя, выполняющую респираторную функцию, от засорения. Когда Aphrodita зарыта в грунт, наружу торчат лишь спинные усики нескольких самых задних сегментов. Крупные черви Polyodontidae (рис. 9, 7), строящие собственные толстые илистые трубки, имеют параподии, слабо расчлененные на дорсальную и вентральную ветви, но снабженные специальными «паутинными железами», секрет которых в виде тонких золотистых волокон служит основой для стенок их трубок; эти золотистые волокна образуются хетобластами, а потому их следует рассматривать как одну из модификаций щетинок. Наиболее сложно устроенные параподии с большим набором различных щетинок наблюдаются у Sigalionidae (рис. 9, 6) и Eulepethidae (рис. 9, 8), что связано с закапыванием в грунт и обитанием в трубках. Представители Chrysopetalidae имеют параподии (рис. 9, 9), на спинной ветви которых в виде широкого веера располагаются золотистые крупные палены, прикрывающие наподобие элитр спинную сторону червя.
Один из наиболее обычных и простых способов передвижения чешуйчатых червей — «марширование» (шагательные движения), при котором невропо-диальные щетинки действуют как рычаги, все тело животного как бы передвигается на ходулях. Довольно сложная работа самих параподии, как показали недавние экспериментальные исследования, строго координируется туловищным мозгом.
Нам пришлось наблюдать движения Lepidonotus squamatus в аквариуме: при спокойном состоянии животного тело его неподвижно, однако параподии все время поочередно выдвигаются наружу, а затем сокращаются — втягиваются обратно, что представляет собой чисто дыхательные движения. Когда животное потревожено, тело производит легкие волновые изгибы, начинающиеся от заднего конца, а параподии резко усиливают свою деятельность: во втянутом состоянии они выносятся вперед, а затем в расправленном виде с сильно выдвинутыми щетинками производят отталкивающий удар назад наподобие движения весла. При поступательном движении червя спинные его усики и головные щупальца обычно неподвижны. У всех чешуйчатых червей при поступательном движении их форма и размеры меняются незначительно.
Остановимся несколько подробнее на поисковых движениях Iphione muri-cata, обитающего в прибрежной зоне Баренцева моря. Днем эти черви сидят неподвижно, плотно прикрепившись к камням, причем если их потревожить, они еще плотнее прижимаются к субстрату, от которого их можно оторвать лишь с большим трудом. Двигаться в поисках пищи они начинают через несколько часов после захода солнца, а перед восходом солнца сразу останавливаются. За ночь они успевают передвинуться всего на несколько сантиметров, причем некоторые из них к утру возвращаются на прежнее место. При ночных путешествиях элитры ритмично подымаются и опускаются, пропуская под собой ток воды. Параподии приступают к работе с заднего конца тела, одна волна движений параподии длится около 5 секунд, после чего наступает пауза в 2 секунды; параподии на каждом сегменте работают в противоположном направлении. Иногда вместо поступательного движения у Iphione наблюдается верчение на одном месте.
0 комментариев