Навигация
Проникновение вируса в клетку
23317
знаков
0
таблиц
0
изображений
3. Проникновение вируса в клетку
Адсорбированные вирусы проникают в клетку с помощью эндоцитоза или путем слияния с клеточной мембраной. Оказавшись в цитоплазме, вирусы освобождаются от большинства белков (раздевание) и начинают реплицироваться. Проникновение в клетку, раздевание и репродукция вирусов зависят от интенсивности энергетического метаболизма клетки и биохимических изменений, происходящих в клеточной мембране и цитоскелете. Так, при температуре ниже 37*С проникновение вирусов в клетку замедляется. Пусковым фактором проникновения обычно служит связывание некоторых поверхностных белков вируса с мембранными рецепторами клетки. Эти белки представлены на поверхности вирусов по крайней мере несколькими молекулами, а количество мембранных рецепторов обычно достигает нескольких сотен. В месте контакта вируса с клеточной мембраной происходит агрегация рецепторов, которая запускает механизм внутриклеточной передачи сигнала и стимулирует изменения в клеточной мембране. Адсорбция вируса обычно воспринимается клеткой как присоединение "нормального" лиганда к соответствующему рецептору.
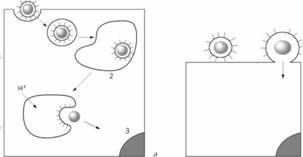
Адсорбция многих вирусов запускает эндоцитоз, начинающийся с образования на мембране окаймленных ямок, покрытых клатрином. Затем формируются эндосомы, в составе которых вирусы поступают в цитоплазму. Данный способ проникновения в клетку характерен для пикорнавирусов, вирусов гриппа и аденовирусов. Последующее слияние вирусов с мембраной эндосом стимулируется понижением рН в эндосоме.
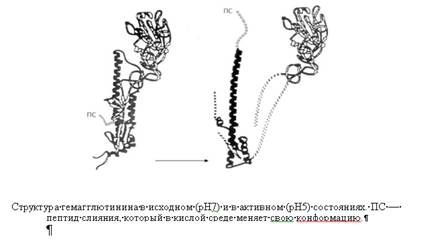
Влияние рН на процесс проникновения хорошо изучено у вируса гриппа. В адсорбции этих вирусов, агрегации рецепторов и эндоцитозе важную роль играют гемагглютинины внешней оболочки. Конформационные изменения гемагглютинина, возникающие при низком рН в эндосоме, приводят к выходу на поверхность молекулы амфифильных доменов, что приводит к слиянию внешней оболочки вируса и эндосомальной мембраны.
На молекулярном уровне процессы слияния с мембраной и раздевания большинства вирусов изучены плохо. В результате слияния липиды и белки внешней оболочки вируса смешиваются с липидами и белками клеточной мембраны, а нуклеокапсид оказывается в цитоплазме. У сложных вирусов в адсорбции и слиянии с клеточной мембраной могут последовательно участвовать разные белки внешней оболочки.
Есть данные, что в разных тканях или на разных поверхностях эпителиальных клеток механизмы адсорбции вирусов и их проникновения в клетку неодинаковы.
4. Синтез вирусных белков
В основе этого синтеза лежит тот же механизм, что и при синтезе белка в нормальных клетках. У РНК-содержащих пикорнавирусов функцию иРНК выполняют плюс-нити. У них односпиральная вирионная РНК транслируется с образованием одного гигантского полипептида, который затем расщепляется на отдельные функциональные белки. В синтезе полновирусного белка выражена постоянная во времени трансляция всех генов вирусной РНК. У орто-, рабдо- и парамиксовнрусов вирионная односпиральная РНК не транслируется, а транслируется комплементарная ей плюс-нить, поэтому синтез вирусных специфических белков начинается после образования вирусной иРНК (плюс-нити), которая комплементарна вирионной РНК-Минус-нити синтезируются на плюс-нити вирионной РНК-зависимой полимеразой (РНК-транскриптазой), находящейся в составе вириона в качестве структурного компонента. Синтезирующиеся вирионной полимеразой иРНК являются моноцистронными и значительно короче вирионной РНК. В процессе вирусной инфекции происходят распад клеточных полисом и образование вирусспецифических полисом.
Синтез вирусспецифического белка зависит от синтеза вирусной иРНК, но и влияет на него: если синтез белка нарушен, происходит затоваривание вновь образующейся иРНК в местах ее синтеза и тормозится дальнейший ее синтез.
Вирусные белки в процессе инфекции синтезируются в избыточном количестве, чем требуется для образования инфекционного вируса. Например, в клетках, инфицированных вирусами герпеса, в вирусное потомство включается только около 35% от общей массы вирусспецифических белков, синтезированных в клетках.
У большинства вирусов синтез белков осуществляется в цитоплазме; относительно ядерной локализации синтеза белков некоторых вирусов существует сомнение. Известно, что вирусные белки могут синтезироваться в одних структурах, а накапливаться— в других. Механизмы, ответственные за миграцию вирусных белков в ядро, не выяснены. Известно лишь, что отсутствие аргинина в среде приводит к подавлению миграции структурных белков вируса герпеса от места их синтеза (цитоплазмы) к месту сборки вирионов (ядру), хотя синтез ДНК и белка вируса не нарушен.
На разных стадиях инфекционного цикла могут преимущественно образовываться то одни, то другие группы вирусспецифических белков. Скорость их регулируется либо на уровне транскрипции (с образованием иРНК), либо на уровне трансляции (считывание иРНК на рибосомах).
В зараженной клетке непропорционально накапливаются иРНК с разных вирусных генов. Механизм этой непропорциональности заложен в самой вирусной частице. Этот же механизм определяет разную эффективность образования различ- ых белков. Стандартная вирусная частица содержит одну молекулу РНК и до 10 тыс. молекул белков. Помимо структур-ных белков, в зараженной клетке могут синтезироваться и не-структурные (но кодируемые вирусной РНК) белки. Наряду с синтезом белков в клетке при репродукции вируса гриппа происходит синтез и углеводных цепей, входящих в состав гликопротеидов. Присоединение углеводов осуществляется с помощью трансфераз, которые являются клеточными ферментами. Синтез липидов также осуществляется клеткой. Вирусная оболочка формируется при включении липидов из плазматической мембраны клетки-хозяина. Синтез вирусных нуклеиновых кислот и вирусспецифических белков происходит почти одновременно и не менее чем на 1 ч опережает созревание вирусных частиц.
Похожие работы
... , вирус Эбола и другие, способны ассоциироваться с "плотами" клеточной мембраны. Также филовирусы Эбола и Марбурга проникают в клетки путем кавеолин-опосредованного эндоцитоза, при котором белок кавеолин связан с микродоменами плазмалеммы. Механизм проникновения вирусов путем эндоцитоза, независимого от клатрина и кавеолина и опосредованного липидными площадками плазмалеммы - "плотами", главным ...
... . У штаммов HIV с мутациями, инактивирующими vif, вирионы имеют нормальный вид, содержат полный набор РНК и ферментов, но заражают клетки намного менее эффективно. 3. Этапы заражения клетки вирусом СПИДа Первый шаг любой вирусной инфекции - связывание вирусной частицы с компонентом мембраны заражаемой клетки. Для HIV роль такого рецепторного компонента играет белок, называемый антигеном CD4.( ...
... фагами, либо путем выталкивания участков цитоплазмы, либо, наконец, путем выхода отдельных вирионов или небольших их групп. Некоторые вирусы животных с трудом освобождаются из клеток в культурах in vitro; в живом организме выходу таких вирусов из клеток и их распространению способствует захват поврежденных вирусом клеток фагоцитами и их переваривание. Вирусы растений обычно не освобождаются путем ...
... ) выделяют конъюгативные и неконъюгативные плазмиды. Плазмиды могут встраиваться в хромосому бактерий- интегративные плазмиды или находиться в виде отдельной структуры- автономные плазмиды ( эписомы). Классификация и биологическая роль плазмид. Функциональная классификация плазмид основана на свойствах, которыми они наделяют бактерии. Среди них- способность продуцировать экзотоксины и ферменты, ...
0 комментариев