Центральный механизм регуляции дыхания. Дыхательный центр. Опишите путь, проходимый молекулой кислорода при диффузии из альвеолы в эритроциты
Функциональные связи гипоталамуса с гипофизом. Роль гипофиза в регуляции деятельности эндокринных желез. Опишите регуляцию функций яичек у мужчин
Проблема локализации функций в коре большого мозга
Навигация
Центральный механизм регуляции дыхания. Дыхательный центр. Опишите путь, проходимый молекулой кислорода при диффузии из альвеолы в эритроциты
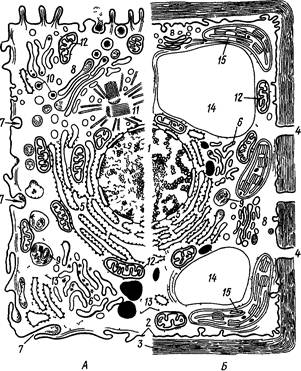
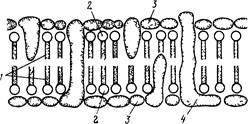
Строение и основные свойства клеточных мембран
45125
знаков
0
таблиц
0
изображений
2. Центральный механизм регуляции дыхания. Дыхательный центр. Опишите путь, проходимый молекулой кислорода при диффузии из альвеолы в эритроциты.
Дыхание регулируется нервным и гуморальным путем. Дыхательный центр находится в продолговатом мозге, содержит два отдела: центр вдоха и центр выдоха. При выдохе легкие спадаются и возбуждаются рецепторы, находящиеся в альвеолах. Возникшие возбуждения передаются по нервам в дыхательный центр, в отдел вдоха. Этот отдел приходит в возбужденное состояние и посылает импульсы в спинной мозг, а оттуда импульсы поступают к наружным межреберным мышцам и диафрагме. Они сокращаются и вызывают расширение грудной клетки - происходит вдох. Теперь отдел вдох не получает раздражения и не посылает возбуждения. Дыхательные мышцы расслабляются, грудная клетка спадает и происходит выдох. Вдох порождает выдох.
Процесс дыхания слагается из трех основных фаз: внешнее дыхание (обмен газов между внешней средой и кровью), транспорт газов кровью и внутреннее (тканевое) дыхание (обмен газов между кровью и тканями). Внешнее дыхание, функцию которого выполняют органы дыхательной системы, осуществляется различными способами.
Согласованность, ритмичность сокращений и расслаблений дыхательных мышц обусловлены поступающими к ним по нервам импульсами от дыхательного центра продолговатого мозга. И.М. Сеченов в 1882 г. установил, что примерно через каждые 4 сек, в дыхательном центре автоматически возникают возбуждения, обеспечивающие чередование вдоха и выдоха. Дыхательный центр изменяет глубину и частоту дыхательных движений, обеспечивая оптимальное содержание газов в крови. Гуморальная регуляция дыхания состоит в том, что повышение концентрации углекислого газа в крови возбуждает дыхательный центр - частота и глубина дыхания увеличиваются, а уменьшение СО2 понижает возбудимость дыхательного центра - частота и глубина дыхания уменьшаются.
Дыхательный центр
Под дыхательным центром следует понимать совокупность нейронов специфических (дыхательных) ядер продолговатого мозга, способных генерировать дыхательный ритм.
В нормальных (физиологических) условиях дыхательный центр получает афферентные сигналы от периферических и центральных хеморецепторов, сигнализирующих соответственно о парциальном давлении О2 в крови и концентрации Н+ во внеклеточной жидкости мозга. В период бодрствования деятельность дыхательного центра регулируется дополнительными сигналами, исходящими из различных структур ЦНС. У человека это, например, структуры, обеспечивающие речь. Речь (пение) может в значительной степени отклонить от нормального уровень газов крови, даже снизить реакцию дыхательного центра на гипоксию или гиперкапнию. Афферентные сигналы от хеморецепторов тесно взаимодействуют с другими афферентными стимулами дыхательного центра, но, в конечном счете, химический, или гуморальный, контроль дыхания всегда доминирует над нейрогенным. Например, человек произвольно не может бесконечно долго задерживать дыхание из-за нарастающих во время остановки дыхания гипоксии и гиперкапнии.
Дыхательный центр выполняет две основные функции в системе дыхания: моторную, или двигательную, которая проявляется в виде сокращения дыхательных мышц, и гомеостатическую, связанную с изменением характера дыхания при сдвигах содержания О2 и СО2 во внутренней среде организма.
Двигательная функция дыхательного центра заключается в генерации дыхательного ритма и его паттерна. Под генерацией дыхательного ритма понимают генерацию дыхательным центром вдоха и его прекращение (переход в экспирацию). Под паттерном дыхания следует понимать длительность вдоха и выдоха, величину дыхательного объема, минутного объема дыхания. Моторная функция дыхательного центра адаптирует дыхание к метаболическим потребностям организма, приспосабливает дыхание в поведенческих реакциях (поза, бег и др.), а также осуществляет интеграцию дыхания с другими функциями ЦНС.
Гомеостатическая функция дыхательного центра поддерживает нормальные величины дыхательных газов (O2, CO2) и рН в крови и внеклеточной жидкости мозга, регулирует дыхание при изменении температуры тела, адаптирует дыхательную функцию к условиям измененной газовой среды, например при пониженном и повышенном барометрическом давлении.
В организме газообмен О2 и СО2 через альвеолярно-капиллярную мембрану происходит с помощью диффузии. Диффузия О2 и СО2 через аэрогематический барьер зависит от следующих факторов: вентиляции дыхательных путей; смешивания и диффузии газов в альвеолярных протоках и альвеолах; смешивания и диффузии газов через аэрогематический барьер, мембрану эритроцитов и плазму альвеолярных капилляров; химической реакции газов с различными компонентами крови, и, наконец, от перфузии кровью легочных капилляров.
Диффузия газов через альвеолярно-капиллярную мембрану легких осуществляется в два этапа. На первом этапе диффузионный перенос газов происходит по концентрационному градиенту через тонкий аэрогематический барьер, на втором — происходит связывание газов в крови легочных капилляров, объем которой составляет 80—150 мл, при толщине слоя крови в капиллярах всего 5—8 мкм и скорости кровотока около 0,1 мм*с-1. После преодоления аэрогематического барьера газы диффундируют через плазму крови в эритроциты.
Значительным препятствием на пути диффузии О2 является мембрана эритроцитов. Плазма крови практически не препятствует диффузии газов в отличие от альвеолярно-капиллярной мембраны и мембраны эритроцитов
3.Нефрон – структурно-функциональная единица почки. Образование клубочкового фильтра.
В каждой почке у человека содержится около 1 млн функциональных единиц — нефронов, в которых происходит образование мочи. Каждый нефрон начинается почечным, или мальпигиевым, тельцем — двустенной капсулой клубочка (капсула Шумлянского—Боумена), внутри которой находится клубочек капилляров. Внутренняя поверхность капсулы выстлана эпителиальными клетками; образующаяся полость между висцеральным и париетальным листками капсулы переходит в просвет проксимального извитого канальца. Особенностью клеток этого канальца является наличие щеточной каемки — большого количества микроворсинок, обращенных в просвет канальца. Следующий отдел нефрона — тонкая нисходящая часть петли нефрона (петли Генле). Ее стенка образована низкими, плоскими эпителиальными клетками. Нисходящая часть петли может опускаться глубоко в мозговое вещество, где каналец изгибается на 180°, и поворачивает в сторону коркового вещества почки, образуя восходящую часть петли нефрона. Она может включать тонкую и всегда имеет толстую восходящую часть, которая поднимается до уровня клубочка своего же нефрона, где начинается дистальный извитой каналец. Этот отдел канальца обязательно прикасается к клубочку между приносящей и выносящей артериолами в области плотного пятна. Клетки толстого восходящего отдела петли Генле и дистального извитого канальца лишены щеточной каемки, в них много митохондрий и увеличена поверхность базальной плазматической мембраны за счет складчатости. Конечный отдел нефрона — короткий связующий каналец, впадает в собирательную трубку. Начинаясь в корковом веществе почки, собирательные трубки проходят через мозговое вещество и открываются в полость почечной лоханки. Диаметр капсулы клубочка около 0,2 мм, общая длина канальцев одного нефрона достигает 35—50 мм. Исходя из особенностей структуры и функции почечных канальцев, различают следующие сегменты нефрона: 1) проксимальный, в состав которого входят извитая и прямая части проксимального канальца; 2) тонкий отдел петли нефрона, включающий нисходящую и тонкую восходящую части петли; 3) дистальный сегмент, образованный толстым восходящим отделом петли нефрона, дистальным извитым канальцем и связующим отделом. Канальцы нефрона соединены с собирательными трубками: в процессе эмбриогенеза они развиваются самостоятельно, но в сформировавшейся почке собирательные трубки функционально близки дистальному сегменту нефрона.
В почке функционирует несколько типов нефронов: суперфициальные (поверхностные), интракортикальные и юкстамедуллярные. Различие между ними заключается в локализации в почке, величине клубочков (юкстамедуллярные крупнее суперфициальных), глубине расположения клубочков и проксимальных канальцев в корковом веществе почки (клубочки юкстамедуллярных нефронов лежат у границы коркового и мозгового вещества) и в длине отдельных участков нефрона, особенно петель нефрона. Суперфициальные нефроны имеют короткие петли, юкстамедуллярные, напротив, длинные, спускающиеся во внутреннее мозговое вещество почки. Характерна строгая зональность распределения канальцев внутри почки
Большое функциональное значение имеет зона почки, в которой расположен каналец, независимо от того, находится ли он в корковом или мозговом веществе. В корковом веществе находятся почечные клубочки, проксимальные и дистальные отделы канальцев, связующие отделы. В наружной полоске наружного мозгового вещества находятся нисходящие и толстые восходящие отделы петель нефронов, собирательные трубки; во внутреннем мозговом веществе располагаются тонкие отделы петель нефронов и собирательные трубки. Расположение каждой из частей нефрона в почке чрезвычайно важно и определяет форму участия тех или иных нефронов в деятельности почки, в частности в осмотическом концентрировании мочи.
Клубочковая фильтрация
Мысль о фильтрации воды и растворенных веществ как первом этапе мочеобразования была высказана в 1842 г. немецким физиологом К. Людвигом. В 20-х годах XX столетия американскому физиологу А. Ричардсу в прямом эксперименте удалось подтвердить это предположение — с помощью микроманипулятора пунктировать микропипеткой клубочковую капсулу и извлечь из нее жидкость, действительно оказавшуюся ультрафильтратом плазмы крови.
Ультрафильтрация воды и низкомолекулярных компонентов из плазмы крови происходит через клубочковый фильтр. Этот фильтрационный барьер почти непроницаем для высокомолекулярных веществ. Процесс ультрафильтрации обусловлен разностью между гидростатическим давлением крови, гидростатическим давлением в капсуле клубочка и онкотическим давлением белков плазмы крови. Общая поверхность капилляров клубочка больше общей поверхности тела человека и достигает 1,5 м2 на 100 г массы почки. Фильтрующая мембрана (фильтрационный барьер), через которую проходит жидкость из просвета капилляра в полость капсулы клубочка, состоит из трех слоев: эндотелиальных клеток капилляров, базальной мембраны и эпителиальных клеток висцерального (внутреннего) листка капсулы— подоцитов.
Клетки эндотелия, кроме области ядра, очень истончены, толщина цитоплазмы боковых частей клетки менее 50 нм; в цитоплазме имеются круглые или овальные отверстия (поры) размером 50—100 нм, которые занимают до 30 % поверхности клетки. При нормальном кровотоке наиболее крупные белковые молекулы образуют барьерный слой на поверхности пор эндотелия и затрудняют движение через них альбуминов, ограничивая тем самым прохождение форменных элементов крови и белков через эндотелий. Другие компоненты плазмы крови и вода могут свободно достигать базальной мембраны.
Базальная мембрана является одной из важнейших составных частей фильтрующей мембраны клубочка. У человека толщина базальной мембраны 250—400 нм. Эта мембрана состоит из трех слоев — центрального и двух периферических. Поры в базальной мембране препятствуют прохождению молекул диаметром больше 6 нм.
Наконец, важную роль в определении размера фильтруемых веществ играют щелевые мембраны между «ножками» подоцитов. Эти эпителиальные клетки обращены в просвет капсулы почечного клубочка и имеют отростки — «ножки», которыми прикрепляются к базальной мембране. Базальная мембрана и щелевые мембраны между этими «ножками» ограничивают фильтрацию веществ, диаметр молекул которых больше 6,4 нм (т. е. не проходят вещества, радиус молекулы которых превышает 3,2 нм). Поэтому в просвет нефрона свободно проникает инулин (радиус молекулы 1,48 нм, молекулярная масса около 5200), может фильтроваться лишь 22 % яичного альбумина (радиус молекулы 2,85 нм, молекулярная масса 43500), 3 % гемоглобина (радиус молекулы 3,25 нм, молекулярная масса 68 000)и меньше 1 % сывороточного альбумина (радиус молекулы 3,55 нм, молекулярная масса 69 000).
Прохождению белков через клубочковый фильтр препятствуют отрицательно заряженные молекулы — полианионы, входящие в состав вещества базальной мембраны, и сиалогликопротеиды в выстилке, лежащей на поверхности подоцитов и между их «ножками». Ограничение для фильтрации белков, имеющих отрицательный заряд, обусловлено размером пор клубочкового фильтра и их электронегативностью. Таким образом, состав клубочкового фильтрата зависит от свойств эпителиального барьера и базальной мембраны. Естественно, размер и свойства пор фильтрационного барьера вариабельны, поэтому в обычных условиях в ультрафильтрате обнаруживаются лишь следы белковых фракций, характерных для плазмы крови. Прохождение достаточно крупных молекул через поры зависит не только от их размера, но и конфигурации молекулы, ее пространственного соответствия форме поры.
Похожие работы

... лизосомы являются производными эндоплазматической сети или аппарата Гольджи. Аппарат Гольджи. Эта структура названа по имени открывшего ее в 1898 г. итальянского ученого К. Гольджи и до недавнего времени признавалась как компонент только животной клетки. Однако с помощью электронного микроскопа выяснилось присутствие аппарата Гольджи почти во всех типах и растительных клеток. Аппарат Гольджи ...
... Формирование бислоя является особым свойством молекул липидов и реализуется даже вне клетки. Важнейшие свойства бислоя: - способность к самосборке - текучесть - ассиметричность. 1.2. Хотя основные свойства биологических мембран определяются свойствами липидного бислоя, но большинство спецефических функций обеспечивается мембранными белками. Большинство из них пронизывают бислой в виде одиночной ...
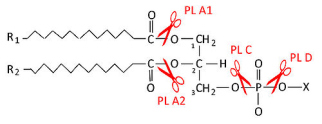
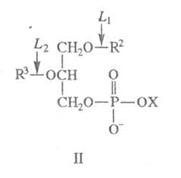
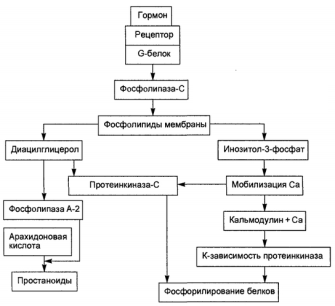

... фосфолипаза относится к ферментам пищеварения и участвует в переваривании липидов пищи. Фосфолипазы яда участвуют в обездвиживании жертвы за счёт лизиса её клеток. 2.2 Классификация и свойства 2.2.1 Цитозольные ФЛА2 История цитозольных фосфолипаз группы А2 началась в 1991 г., когда из цитозоля различных клеток животных был выделен и клонирован белок молекулярной массой 85 кДа, который ...
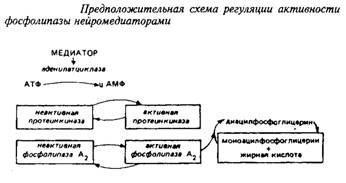

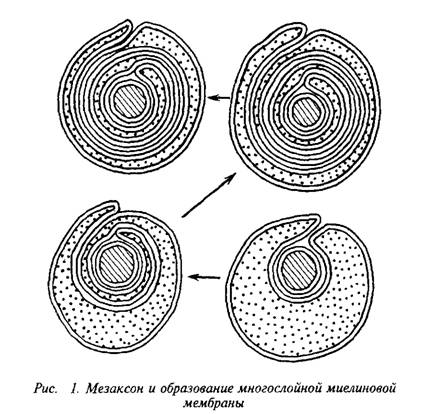
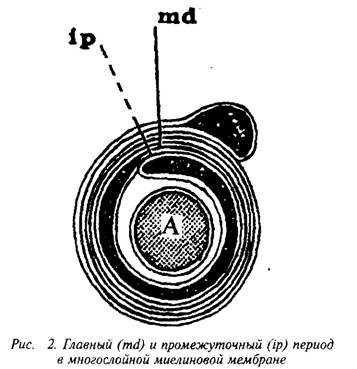
... и инозитолтрифосфат подвергаются химическим превращениям, требующим АТФ и ЦТФ и приводящим к восстановлению три-фосфоинозитида. Таким образом, цикл замыкается и уровень полифосфоинозитидов в мембране восстанавливается. 7. МИЕЛИН В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ Мозг человека содержит 120 г миелина, что составляет одну треть его сухой массы. Миелин – уникальное образование, организация которого ...
0 комментариев