Навигация
ХАРАКТЕРИСТИКИ ОТДЕЛЬНЫХ РЕЦЕПТОРНЫХ СИСТЕМ
90264
знака
4
таблицы
11
изображений
8. ХАРАКТЕРИСТИКИ ОТДЕЛЬНЫХ РЕЦЕПТОРНЫХ СИСТЕМ
Большинство известных в настоящее время нейромедиаторов участвуют в передаче сигнала и через ионотропные, и через метаботропные рецепторы. Это заставляет строить дальнейшее изложение, следуя перечню медиаторов.
Ацетилхолиновые рецепторы. Ацетилхолин как нейромедиатор периферической и центральной нервной системы взаимодействует с двумя видами холинорецепторов: мускариновыми и никотиновыми. Эти подтипы рецепторов отличаются по специфичности взаимодействия с рядом агонистов и антагонистов ацетилхолина. Так, м-ХР избирательно возбуждаются мускарином, а н-ХР.отвечают на аппликацию никотина. Физиологически важным различием между м-ХР и н-ХР является скорость ответа на приходящий сигнал. Считают, что н-ХР предназначены опосредствовать быстрые и непродолжительные эффекты, в то время как м-ХР реагирует более медленно и длительно.
Никотиновые холинорецепторы. н-ХР оказались более изученными биохимически благодаря существованию двух факторов: наличие специфического нейротоксина, способного блокировать функцию рецептора, и обнаружению большого количества этого рецептора в электрических органах рыб. Структура представлена на рис. 7.
Н-ХР содержит 5 субъединиц: две а-субьединицы с Мг = 40 кД, одну р-субъединицу — Мг = 49 кД, одну у-субъединицу — 60 кД и одну 5-субъединицу — Мг = 67 кД. Катионные группы двух молекул ацетилхолина связываются с анионными участками ct-субъединиц. KD взаимодействия АХ с рецептором близко к 10~ М. Открывающийся при контакте с АХ на несколько миллисекунд канал успевает пропустить до 5-Ю ионов К+ и Na+. АХ, диссоциировавший с рецептором, или "избыточный" АХ в синаптической щели быстро расщепляется ферментом ацетилхолинэстеразой, расположенной на постсинаптической мембране в непосредственной близости от рецептора. Ацетилхолинэстераза является одним из самых быстродействующих, высокооборотных ферментов. Таким образом, сигнал резко ограничен во времени. Образовавшийся холин захватывается белками-транспортерами пресинаптической мембраны и служит далее для ресинтеза АХ в терминал и.
Активность рецептора может модулироваться со стороны клетки фосфорилированием отдельных аминокислотных остатков участка, обращенного внутрь клетки. Подвижность рецептора ограничена связью с цитоскелетными белками через так называемый белок 43К.
По характеру влияния веществ на функцию ХР можно выделить: агонистпы, антагонисты и блокаторы. Наиболее известные из них представлены в табл. 3.
Таблица 8Л
Лиганды холинорецепторов
| Типы лигандов | ||
| никотиновые | мускариновые | |
| Агонисты Антагонисты, Блокаторы | Ацетилхолин, карбахол, никотин, лобелии, 1,1 -диметил-4-фенил-пиперазин Гексаметоний, декамстоний а - Бунгаротоксин 6 - Тубокурарин | Ацетилхолин, карбахол, метахолин, мускарин, диметил ацетилхолин Атропин, галл амин, платифиллин |
Способность разных соединений взаимодействовать с этими рецепторами имеет не только теоретическое, но и большое практическое значение. Поскольку нарушение холинергической медиации лежит в основе ряда патогенетических механизмов заболеваний нервной, эндокринной, иммунной систем, то поиск лекарственных веществ, непосредственно воздействующих на пострецепторные механизмы, является эффективным. Действительно, как показала практика, многие используемые в клинике н-холинергические фармпрепараты имеют точкой своего приложения периферические моторные синапсы, ганглии и некоторые хемочувствительные структуры висцеральных систем. Часто в клинике применяют блокаторы н-ХР, среди которых выделяют ганглиоблокаторы и миорелаксанты. Нарушение функции никотиновых ХР лежит в основе тяжелого прогрессирующего заболевания — миастении гравис. Болезнь резко снижает эффективность нервно-мышечных соединений и обусловлена появлением аутоантител к ХР. Показательно, что содержание аутоантител к ХР в крови больных коррелирует с клиническим состоянием: более высокие титры аутоантител наблюдаются у тяжелых больных миастенией. Лечение этой категории больных с помощью иммунодепрессантов вызывает длительные положительные сдвиги в клинической картине заболевания.
Мускариновые рецепторы ацетилхолина. Эта категория рецепторов ацетилхолина относится к категории метаботропных. Общие их характеристики уже описаны выше. Структурная модель м-ХР представлена на рис. 8. Пептидная цепь, образующая основу м-ХР, состоит из 800-950 аминокислотных остатков и связана с углеводными компонентами. Существует большое число подтипов м-ХР, связанных с различными физиологическими эффектами. В частности, рецепторы м2-ХР, локализованные преимущественно в ЦНС и сердце, продуцируют в качестве вторичных мессенджеров цГМФ и подавляют аденилатциклазу. Рецепторы подтипа МрХР, расположенные, в частности, в желудке и симпатических ганглиях, индуцируют образование инозитолфосфатов и диацилглицерола, которые, в свою очередь, ведут к повышению в цитозоле концентрации Са+ и активации протеинкиназы С. Вторичное модулирующее воздействие они оказывают на кальциевые каналы. Различия рецепторов м1 и м2 выражается также в высоком и низком, соответственно, сродстве к ацетилхолину. Последовательность многообразных молекулярных событий, ведущих к ответу клетки на действие лигандов для м-ХР, показана на схеме 1.
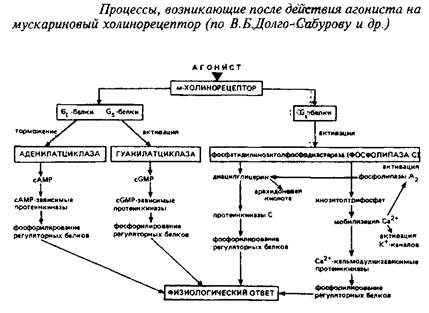
Мускариновые рецепторы АХ связаны с психоэмоциональным восприятием, секрецией слюнных и желудочных желез, функцией сердечно-сосудистой системы и др. Антагонисты м-ХР применяются в клинике в качестве язвазаживляющих, спазмолитических препаратов, а также используются для симптоматического лечения паркинсонизма. В качестве успокаивающих средств, транквилизаторов находят применение в терапии амизил и метамизыл — центральные блсекаторы м-ХР.
Таблица 4.
Специфические агонисты и антагонисты ШМК-рецепторов
| Типы лигандов | Типы ГАМК-рецепторов | |
| ГАМКА | ГАМКВ | |
| Агонисты | Гамма-аминомасляная кислота, мусцимол | .Гамма-аминомасляная кислота, баклофен |
| Антагонисты | Бикукулин, пикротоксин | Дельта-амкно валериановая кислота |
Наконец, мощное влияние на активность холи-нергической передачи оказывают агенты, ингибирующие холи-нестеразу и повышающие тем самым концентрацию АХ в си-наптической щели. Таков механизм действия эзерина и его аналогов, применяемых для снижения внутриглазного давления.
ГАМК-рецепторы. Успехи в идентификации различных типов ГАМК-рецепторов, их биологических и фармакологических характеристик тесно связаны с созданием специфических аго-нистов и антагонистов. По локализации ГАМК-рецепторы подразделяются на центральные и периферические, пре-и постсинаптические. Различают два типа рецепторов ГАМК: бикукулин-чувствительные и баклофен-чувствительные. Наиболее изученным является первый тип рецепторов, который чувствителен также к антагонисту пикротоксинину. Обнаружено, что этот тип рецепторов является быстродействующим и сопряжен с ионными каналами для СР. Другой тип ГАМК-рецепторов относится к медленнодействующим рецепторам, и полагают, что он через G-белок ассоциирован с каналами для ионов К+ и Са+.
Исследования физико-химических свойств очищенного ГАМКд-белка показали, что Мг его находится в пределах 220-270 кди что он представляет собой пентамер гликопротеидли-пидной природы, образующий каналы для ионов хлора.
Особенностью ГАМКА-рецепторов является то, что они содержат специфические участки связывания не только самой ГАМК, но и других физиологически активных соединений. Наиболее интересными и изученными среди них являются лекарственные соединения, объединенные под названием бензо-диазепины а также эндогенные регуляторы пептидной природы — эндозепины.
Среди лекарственных веществ бензодиазепины занимают особое место в связи с их широким лечебным спектром действия: противосудорожного, снотворного, нейротропного, анти-ксиолитического и др.
Важной особенностью функционирования ГАМК-ергической трансмиссии является система удаления выполнившего свою функцию или избыточного лиганда из синаптической щели. В отличие от многих других синаптических систем ГАМК преимущественно претерпевает обратный захват и возвращается в нервные окончания с помощью белков-транспортеров. Они расположены на пресинаптической мембране и несколько похожи по особенностям структуры на метаботропные рецепторы: пептидная цепь, многократно пересекающая мембрану с образованием системы петель над и под мембраной. Белок-транспортер узнает, захватывает и за счет энергии протонного насоса '.переносит ГАМК внутрь терминали.
Места специфического связывания бензодиазепинов находятся на молекулах белка, входящего в структуру рецептора ГАМК. Активация ГАМКА-рецепторов приводит к открытию ионного канала для хлора, а бензодиазепины.при этом удлиняют продолжительность существования открытых ионных каналов, не влияя на их число и скорость транспорта хлора.
Установлено, что участок связывания бензодиазепинов взаимодействует также с эндогенными пептидными регуляторами — эндозепинами. Последние обладают физиологическими эффектами, противоположными бензодиазепинам, — вызывают возбуждение, тревожность и проконфликтное поведение животных. Они подавляют открытие канала для CP, индуцируемое ГАМК, т.е. являются ее эндогенными функциональными антагонистами. Таким образом, бензодиазепины оказались блокаторами участка связывания эвдозепинов, т.е., так сказать, экзогенными антагонистами эндогенных антагонистов ГАМК. Понятно поэтому, что эндозепины обозначают иногда аббревиатурой DBI.
Имеются указания на существование в ЦНС еще одной категории эндогенных антагонистов ГАМК — производных р-кар-болинов. Они также вызывают тревожность, панические состояния у животных и человека.
Что касается второго типа ГАМК-рецетгоров — ГАМКВ, то кроме отмеченных выше особенностей агонистов и антагонистов они характеризуются преимущественно пресинаптической локализацией и сопряженностью с калиевыми, а не с хлор-каналами; локализованы они главным образом.в периферической нервной системе.
Глициновые рецепторы. Радиолигандные исследования позволили локализовать и изучить особенности распределения в центральной нервной системе участков связывания, которые метятся Н-стрихнином. Эти участки, имеющие Кд = 10~ М, являются рецепторами глицина. Наибольшая плотность глициновых рецепторов обнаружена в области ядер.подъязычного и тройничного нервов, локализованных в продолговатом мозге.
Участки связывания стрихнина найдены также в ретикулярных ядрах продолговатого мозга, моста и среднего мозга. Серое вещество спинного мозга также отличается высокой плотностью глициновых рецепторов как в передних, так и в задних рогах.
Глициновый рецептор спинного мозга млекопитающих был очищен с помощью аффинной хроматографии на аминострих-нин-агарозе. Обнаружено, что он представляет собой гликопро-теид-липидный комплекс с Мг = 250 кД, состоящий из 3 полипептидов: 48, 58, 93 кД. Стрихнин и глицин-связывающий сайт расположены на пептиде с Мг - 48 кД, который обладает способностью взаимодействовать с экзогенными лектинами. Встроенный в липосомы белок активирует транспорт ионов ОТ, который блокируется в присутствии стрихнина.
Иммунохимический анализ пептидных компонентов глицинового рецептора с помощью моноклональных антител позволил обнаружить существование общих антигенных детерминант этих рецепторных белков, выделенных из разных объектов: головного и спинного мозга мышей, крыс, свиньи и человека. Более того, интересными являются данные о том, что некоторые участки глицинового и ГАМК-рецепторов иммунологиче-ски идентичны. Этот факт хорошо подтвержден генно-инженерными исследованиями.
До недавнего времени предположение о существовании гомологии между нейрорецепторами I класса, т.е. быстродействующих инотропных рецепторов, выдвигалось лишь в качестве гипотезы. В последние годы одновременно в нескольких лабораториях было показано, что гены рецепторов ГАМК и глицина имеют гомологичные последовательности. Так, оказалось, что имеется примерно 50%-ная гомология между аминокислотными последовательностями а-субъединичной структуры глицинового рецептора с Мг = 48 кД и а- и р-субъединицами ГАМКА-рецептора. Обнаружена 25%-ная гомология между нуклеотидными последовательностями всех трех субъединиц н-ХР.
На рис. 9 представлены консервативные участки известных нейрорецепторов, выявленных при анализе нуклеотидных последовательностей. Характерными особенностями являются высокая степень в гомологии аминокислотной последовательности и расположении трансмембранных участков М1-М4. Обязательное присутствие двух цистеинов в районе 140-150 аминокислоты на расстоянии 14 нуклеотидов друг от друга — отличительная черта нейрорецепторов 1-го класса. Возможно, что все эти нейрорецепторы принадлежат одному семейству белков, кодируемых родственными генами.

Глутаматные рецепторы. Наличие глутаматсвязывающей активности, независимой от присутствия в среде ионов Na, обнаружено практически во всех структурах головного мозга. Наибольшее количество этих участков — в коре больших полушарий, гиппокампе, полосатом теле, среднем мозге и гипоталамусе.
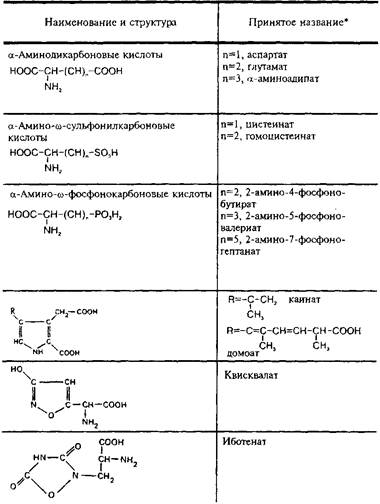
Согласно современным представлениям, существует несколько подтипов глутаматных рецепторов. Их классифицируют прежде всего на основе изучения действия широко известных аналогов глутамата: N-Memn-D-аспартата, а-амино-З-пид-рокси-5-метил-4-изоксазол-пропионовой кислоты, каиновой кислоты, квискваловой кислоты. В табл. 5 представлена структура дикарбоновых возбуждающих аминокислот и некоторых их аналогов. В литературе принято выделять прежде всего два главных подтипа глутаматных рецепторов: NMDA- и не-NMDA-рецепторы. К He-NMDA-рецепторам относятся рецепторы АМРА и каиновой кислоты, сходные по своим физико-химическим свойствам и распространенности в структурах мозга.
Рассмотрим прежде всего NMDA-рецепторы. Они образуют довольно широко распространенный подтип рецепторов глутамата, которые участвуют в разнообразных событиях в ЦНС. В мозге млекопитающих NMDA-связывающие участки локализованы главным образом в кортикальных структурах, базальных ганглиях и сенсорно-ассоциативных системах; наивысшая их плотность обнаружена в гиппокампе. Считают, что они имеют отношение к целому ряду процессов возбуждения, формированию нейрональной пластичности и механизмам памяти, а также к патологическим явлениям нейрональной дегенерации в случае болезни Альцгеймера, церебральной ишемии и др.
Таблица 5
Структура кислых возбуждающих аминокислот и некоторых
их аналогов
NMDA-рецепторы состоят из ряда субъединиц сМг = 40-92 кД и легко олигомеризуются, образуя высокомолекулярные комплексы сМг = 230-270 кД. Эти белки являются гликопротеид-липидными комплексами, формирующими ионные каналы для катионов Na+, К+, Са+. Молекула глугаматного рецептора содержит большое количество гидрофобных аминокислот, которые связаны и с внутренней, и с внешней частью мембраны, организуя взаимодействие с липидами.
Рецептор NMDA имеет несколько участков, взаимодействующих аллостерически. Выделяют пять функционально различных участков, взаимодействие с которыми приводит к изменению активности рецептора:
1) участок связывания нейромедиатора;
2) регуляторный, или коактивирующий, глициновый участок;
3) участок внутри канала, который связывает фенциклидин и родственные соединения;
4) потенциал-зависимый Mg+- связывающий участок;
5) тормозной участок связывания двухвалентных катионов. Наиболее специфический синтетический агонист этих рецепторов — NMDA — не обнаружен в мозге. Предполагается, что кроме глутамата эндогенными медиаторами в этих рецепторах является L-аспартат и L-гомоцистеинат.
Из наиболее известных антагонистов рецепторов NMDA типа можно назвать 0-2-амино-5-фосфоновалериат' и D-2- ами-но-7-фосфоногептаноат. Более специфичны, однако, новые синтетические антагонисты: 3--пропил-Ь-фосфонат и МК-801.СРРл МК-801 - это неконкурентные ингибиторы NMDA, они не действуют непосредственно на участки связывания глутамата.
Своеобразна роль глицинового участка. Глицин в концентрации ОД мкМ увеличивает ответы NMDA-рецептора, и этот эффект не может быть заблокирован стрихнином /напомним, что последний является блокатором самостоятельных глициновых рецепторов). Сам глицин не вызывает ^ответа, а лишь увеличивает частоту открывания канала, не влияя на амплитуду тока при действии агонистов NMDA. Наличке глицина вообще необходимо, поскольку при полном его отсутствии рецептор не активируется L-глутаматом.
Самой важной функцией, которую осуществляет рецептор NMDA в ЦНС, является управление ионным каналом. Важным свойством является способность канала после связывания аго-ниста пропускать ионы Na+ и К+, а также ионы Са+. Предполагают, что внутриклеточный Са+, концентрация которого возрастает при участии рецепторов NMDA, вовлечен в инициацию процессов пластичности развивающегося и взрослого мозга.
Наибольшие токи при активации агонистами возникают при умеренной деполяризации мембраны: от -30 до -20 мВ и уменьшаются при высокой гиперполяризации или деполяризации; следовательно, ионные каналы NMDA-рецепторов являются в определенной мере потенциалзависимыми. Ионы Mg+ селективно блокируют активность рецепторов при таких сдвигах потенциалов. Ионы цинка также ингибируют ответ, но не имеют потенциалзависимого действия, очевидно влияя на другой участок связывания.
К другому подтипу рецепторов глутамата — не NMDA-pe-цепторам — относятся, в частности, рецепторы квискваловой кислоты. Изучение последних привело к пересмотру представления о том, что действие глутамата как нейромедиатора сводится лишь к деполяризации мембраны. Многие типы глутамат-ных рецепторов, и в особенности рецепторы квисквалата, могут функционировать как медленнодействующие метаботропные.
Они вполне соответствуют общим характеристикам метабо-тропных рецепторов, изложенным выше. Пептидная цепочка, составляющая их основу, содержит от 870 до 1000 аминокислотных остатков.
Часть He-NMDA-рецепторов—mGlnRl—реализует сигнал через О0-белки и систему внутриклеточных посредников: инози-толтрифосфатов, диацилглицерола, ионов кальция и др.
Другая разновидность метаботропных He-NMDA-рецепторов — mGlnR2 — реализует сигнал, подавляя синтез цАМФ или активируя синтез цГМФ.
Имеются сведения о том, что рецепторы этой категории участвуют в механизмах синаптогенеза и в изменениях, возникающих при деафферентации. В целом этот тип глутаматных рецепторов, как полагают, участвует в механизмах пластичности аналогично рецепторам NMDA. Но при этом активация рецепторов NMDA блокирует механизм инозитолфосфатной регуляции, связанной с He-NMDA-рецепторами, и наоборот: антагонисты NMDA усиливают действие глутамата на не-NMDA-pe-цепторы.
Весьма интересным примером современных методов изучения рецепторов служит цикл работ с кДНК и ^РНК, кодирующими белки глутаматных рецепторов. Существуют библиотеки полноразмерных генов или их фрагментов мозга млекопитающих. Имея поликлональные антитела к самым разнообразным нейрорецепторам, можно выделить с помощью иммунологического скрининга клоны ДНК, способные продуцировать искомые белковые фракции. Так, недавно из библиотеки кДНК были выделены клоны рекомбинантного фага, дающие положительный иммунологический сигнал на антитела, полученные к глутаматсвязывающему мембранному белку с Мг = 60 кД. Анализ ДНК, выделенной из этого фага, позволил обнаружить наличие вставки кДНК размером 500 нлт, которая способна продуцировать белок с Мг = 14 кД и соответствует узнающей субъединице глутаматного рецептора.-^С помощью этой ДНК была выделена фракция мРНК, комплементарная данной последовательности ДНК. Для доказательства, что выделенная фракция мРНК кодирует синтез глутаматных рецепторов, она была инъецирована в ооциты лягушки, которые являются удобным объектом изучения электрофизиологических свойств нейрорецепторов. Ооциты лягушки обладают эффективным белок-синтезирующим аппаратом, но не имеют собственных нейрорецепторов. После инъекции чужеродной мРНК был измерен мембранный потенциал ооцитов в присутствии глутамата и его аналогов. Оказалось, что выделенная фракция мРНК способна кодировать синтез de novo глутаматных рецепторов каинатного типа.
Возможность одновременного синтеза всех подтипов глутаматных рецепторов в ооцитах лягушки была продемонстрирована другими исследователями. Введение тотальной мРНК, выделенной из мозга крыс, приводило к появлению электрофизиологических ответов у ооцитов на аппликацию NMDA, каината и квисквалата. Более того, ионные токи, регистрируемые на мембране, мало отличались от таковвгх:, обнаруженных на мембранах нейронов. Были, таким офазом, представлены убедительные факты в пользу того, что основные компоненты рецепторного комплекса для глутамата синтезируются совместно, причем биосинтез их не зависит от клетки-носителя и типа мембраны, в которую они затем встраиваются.
Перспективными являются исследования шклада глутаматных рецепторов в патохимию ряда заболеваний ЦНС. Полагают, что эти нейрорецепторы могут служить маркерами деструктив
ных повреждений возбуждающих глутаматергических путей головного мозга и участвовать в аутоиммунных реакциях организма человека. Установление роли глутаматных рецепторов в патогенезе нервно-психических заболеваний — это не единственное направление современной медицины. Появились уже конкретные примеры использования разных антагонистов глутаматных рецепторов против явлений укачивания, токсического действия высоких парциальных давлений кислорода, при лечении инсультов и др.. Кроме того, антагонисты глутаматных рецепторов могут составить основу для создания малотоксичных инсектицидных препаратов для сельского хозяйства.
Адренорецепторы История изучения адренорецепторов тесно связана с открытием биологической функции катехолами-нов в клетках надпочечников. Гипотеза о существовании этого вида рецепторов в самых разнообразных клетках наряду с исследованиями ХР оказалась наиболее плодотворной для развития теории взаимодействия физиологически активных веществ с рецепторами. Несмотря на то что адренорецепторы ъ нервной ткани присутствуют в относительно небольшом количестве, они играют важную роль в регуляции психоэмоциональных функций и деятельности всех отделов сердечно-сосудистой системы.
Адренорецепторы подразделяют на два типа:![]() , в зависимости от связывания адренотропных лигандов. Адренорецепторы
, в зависимости от связывания адренотропных лигандов. Адренорецепторы![]() и
и![]() , в свою очередь, могут быть разделены на подклассы
, в свою очередь, могут быть разделены на подклассы ![]() и
и![]() ,
,![]() и
и![]()
Для![]() ,
,![]() ,
,![]() и
и![]() -рецепторов характерна преимущественно постсинаптическая локализация, а для а2-,, вторичный мессенд-жер, служащий активатором опять-таки протеинкиназ, но другого типа, нежели протеинкиназы, активируемые в результате включения aj-рецепторов.
-рецепторов характерна преимущественно постсинаптическая локализация, а для а2-,, вторичный мессенд-жер, служащий активатором опять-таки протеинкиназ, но другого типа, нежели протеинкиназы, активируемые в результате включения aj-рецепторов.
G-белки — передатчики и модификаторы сигнала от собственного рецепторного белка — также имеют существенные отличия в разных классах адренергических рецепторов. Важным общим их свойством является взаимодействие х ГТФ и ГДФ. В неактивном состоянии они связаны с ГДФ. При взаимодействии нейромедиатора с рецепторным белком последний вступает в контакт с G-белком и меняет его конформацию так, что на место ГДФ становится ГТФ. С этого момента G-белок приобретает способность воздействовать на активность белка-эффектора. Однако G-белок является ГТФазой, быспро расщепляющей ГТФ до ГДФ. В результате он переходит в исходное состояние.
Заметим, что G-белок а2-адренорецетхиэтав, обозначаемый Gs, активирует аденилатциклазу, обеспечивая, >в конечном счете, повышение уровня цАМФ и, далее, стимуляцию протеин-киназ. G-белки![]() - и
- и![]() -адренорецепторов — G-белки —обеспечивают, напротив, подавление активности щиклазы, снижение уровня цАМФ и, соответственно, снижение активности определенных протеинкиназ.
-адренорецепторов — G-белки —обеспечивают, напротив, подавление активности щиклазы, снижение уровня цАМФ и, соответственно, снижение активности определенных протеинкиназ.
Собственно рецепторный белок представляет собой пептидную цепочку с Мг порядка 60-80 кД, пронизывающую внешнюю мембрану так, что ее С-конец и ряд петлеобразных участков экспонированы наружу, а N-конец и опять-таки ряд петлеобразных участков обращены внутрь клетки. Нейромедиатор, взаимодействуя с обращенными наружу участками цепи, меняет конформацию всего белка, так что участки, обращенные внутрь, приобретают сродство к Gg-белку, Gj-белку или к О0-белку. "Выбор" одного из этих белков определяется тонкими особенностями структуры R-бел-ка в области, обращенной внутрь клетки. Заметим также, что участки пептидной цепи, содержат по 20-25 преимущественно гидрофобных аминокислот, а петлевидные участки вне мембраны состоят преимущественно из гидрофильных аминокислот. N-конец полипептида обычно яддикозилирован, а на С-конце есть остатки серина и треонина, wropbie могут фосфорилироваться; последний процесс модулирует активность рецептора.
Практически любое изменение функции![]() и
и![]() -адренорецепторов сопровождается активацией системы внутриклеточных посредников, которые способны избирательно передавать внешний сигнал в цитоплазму и на генетический аппарат клетки. В этом случае геном нейрона может регулировать биосинтез мембранных компонентов или активировать процессы, связанные с их фосфорилированием. Последние реакции приводят к изменению хемочувствительности нервных клеток, иными словами, изменению "информационной емкости" нейронов, и их связывают с механизмом запоминания.
-адренорецепторов сопровождается активацией системы внутриклеточных посредников, которые способны избирательно передавать внешний сигнал в цитоплазму и на генетический аппарат клетки. В этом случае геном нейрона может регулировать биосинтез мембранных компонентов или активировать процессы, связанные с их фосфорилированием. Последние реакции приводят к изменению хемочувствительности нервных клеток, иными словами, изменению "информационной емкости" нейронов, и их связывают с механизмом запоминания.
Дофаминовые рецепторы. Прогресс в изучении структуры и функции дофаминовых рецепторов прежде всего был связан с обнаружением их антагонистов: производных фенотиазепама, галоперидола, спиропервдола и др. Радиолигандные исследования с помощью этих соединений показали, что в головном мозге имеются участки специфического связывания Н-галоперидола, Н-спироперидола и Н-апоморфина с параметрами, колеблющимися в пределах Кд ~7 нМ и Вмакс — 240-400 пмоль/мг белка.
Гетерогенность дофаминовых рецепторов ЦНС подтверждается не только биохимическими и фармакологическими экспериментами in vitro. Анализ влияния различных дофаминсодер-жащих препаратов на поведение крыс обнаружил разные стереотипные двигательные реакции. Оказалось, что вращение крыс в ту или иную сторону опосредуется двумя разными классами дофаминовых рецепторов.
Все дофаминергические рецепторы являются метаботропны-ми, сопряженными с аденилатциклазой. Они классифицируются на 4 типа: Д^, Д2-, Д3 - и Д4 по параметрам связывания их с агонистами и антагонистами и по белковой системе, трансформирующей сигнал. Наиболее изучены Др и Д2-рецепторы. Д j -рецепторы активируют аденилатциклазу через 6£-белок, а Д2-рецепторы, напротив, подавляют аденилатциклазу через Gs-белок. Первые подавляются лишь относительно высокими, микромолярными концентрациями нейролептиков, в то время как для подавления вторых достаточны наномолярные концентрации. Количество и распространение Дри Д2-рецепторов выше, чем Д3 и Д4 в ткани мозга, Д2-рецепторы локализованы преимущественно в клетках ниг-ростриарной и мезкортиколимбической систем. Рвотное действие апоморфина — результат его взаимодействия с Д2-рецепто-рами. Именно нарушение, несбалансированное усиление деятельности систем, связанных с Д2-рецепторами, ассоциируют с синдромом шизофрении. Подавление Дг- и Д2-рецепторов индуцирует нарушения различных форм стереотипных двигательных реакций. Одно из их проявлений — болезнь Паркинсона.
Рецепторы серотонина. Описание рецепторов серотонина облегчается их сходством с рецепторами норадреналина и дофамина. Это метаботропные рецепторы, локализованные как в мозге, так и на периферии. Известно несколько подтипов рецепторов серотонина, в частности 5НТ\, сопряженный с аде-нилатциклазой, и 5НТ2, сопряженный с фосфолипазой С. Последний особенно распространен в постсинаптических мембранах мозга. Специфические антагонисты — спиперон и кетансерин. Сложным является отношение этих рецепторов с производными лизергиновой кислоты, в том числе LSD. По отношению к некоторым разновидностям 5НТ2-рецепторов он является агонистом, а к другим — антагонистом.
Активация 5НТ2 входит в систему реакций, индуцирующих ортодоксальный сон. По-видимому, дефицит или ингибирова-ние 5НТ2 связано с депрессивными состояниями, а активация некоторых из них — с галлюцинациями. Нарушения серотонин-ергической системы связывают и с такой патологией, как тяжелые формы мигрени. Существует характерный двигательный синдром, регулируемый 5НТ2-рецепторами, — "отряхивание мокрой собаки", используемый нередко в исследованиях серотонинергической системы.
Функции 5НТ2-рецепторов, также представленных и в мозге, и на периферии, изучены в меньшей мере. Одна из причин этого — отсутствие высокоспецифичных антагонистов.
Многообразными являются периферические эффекты активации рецепторов серотонина всех подтипов. Главные из них состоят в сокращении сосудов, положительных хроно- и инотропных воздействиях на сердце, а также в стимуляции аггрегации тромбоцитов.
Рецепторы гистамина. К настоящему времени выявлены и исследованы три типа рецепторов гистамина: Hj и Н? — постсинаптические и Н3 - пресинаптические. Рецепторы Hi являются метаботропными, они сопряжены с фосфолипазой С и индуцируют образование инозитол-трифосфата и диацилглицерола. Ингибиторами Я^рецепторов служат димедрол, фенкарол, супрастин и другие соединения, получившие широкое применение в медицине. Метаботропными являются также рецепторы Н2, однако они сопряжены с аденилатциклазой и индуцируют повышение уровня циклического АМФ. Хорошо изученным ингибитором Н2-рецепторов является циметидин — известный противоязвенный агент. Пресинаптические рецепторы гистамина Н3 являются ауторецепторами. Они подавляют выход гистамина из нервных окончаний, т.е. осуществляют обратную связь между уровнем гистамина в синапсе и его секрецией.
Рецепторы пуринов. Последнее десятилетие ознаменовалось накоплением данных об обширном семействе рецепторов разнообразных пуринов: аденозина, АТФ, относительно мало изученного макроэргического соединения диаденозинтетрафосфа-та, а также АМФ и АДФ. Рецепторы пуринов делят на две большие группы: рецепторы, преимущественно взаимодействующие с аденозином, и рецепторы, преимущественно взаимодействующие с АТФ и АФА. Рецепторы аденозина Р1 являются медленными метаботропными рецепторами. Среди них есть рецепторы, подавляющие системы синтеза цАМФ, и рецепторы, напротив, активирующие синтез цАМФ.
Особого рассмотрения заслуживают свойства рецепторов А1. Значительная их часть локализована на пресинаптических мембранах. Довольно специфическими ингибиторами рецепторов А1 являются теофилин и кофеин. Иначе говоря, введение этих широко известных психостимулирующих агентов ведет к увеличению уровня цАМФ в некоторых постсинаптических структурах. Заметим, что кофеин и теофилин являются также довольно специфичными ингибиторами фосфодиэстераз, что тоже способствует повышению уровня цАМФ. Такое двустороннее действие этих соединений на уровень цАМФ позволяет понять их особое значение как психостимуляторов.
Рецепторы А1 включаются не только аденозином, но и АМФ, причем последний может происходить не только из нервных окончаний, но и из других образований мозга. Истощение энергетических систем мозга, связанное с образованием АМФ, может служить, таким образом, сигналом для включения рецепторов А1. Это позволяет лучше понять ряд физиологических эффектов, которые наблюдаются при срабатывании рецепторов А1: успокаивающие, седативные, противосудорожные. Иначе говоря, эти рецепторы выступают как защитники энергетических резервов мозга в экстремальных ситуациях. С этим же сопряжены их гипотензивные эффекты. Из периферических реакций на включение рецептора А1 отметим брадикардию и воздействие на автономные проводящие системы сердца. Адено-зин и его производные оказались перспективными средствами для лечения аритмий.
Из общих физиологических эффектов аденозиновых рецепторов А2 отметим стимуляцию глюконеогенеза и подавление агрегации тромбоцитов.
Рецепторы Р2, имеющие наибольшее сродство к АТФ и АФ4А и наименьшее к аденозину и АМФ, также делятся на несколько подтипов. В отличие от аденозиновых рецепторов среди них есть быстрые, канальные рецепторы заслуживает периферический эффект такого макроэргического соединения, как АДФ —стимуляция тромбоцитов.
Рецепторы нейропептидов. Все рецепторы нейропептидов являются метаботропными, медленными рецепторами. Наиболее изучены опиатные рецепторы.
Опиаты — морфин и родственные ему соединения — известны в медицине с древних времен. Их применяли, как правило, в качестве обезболивающих и наркотических средств, способных влиять на психику человека. Однако хэпиатные рецепторы, которые являются объектами действия опиатов в ЦНС, открыты сравнительно недавно, в начале 70-х годов. Фармакологическими исследованиями было показано, что ОР связывают большое число синтетических и природных лигандов. Первыми изученными лигандами ОР были экзогенные вещества — морфин, дигидроморфин, норморфин, леворфанол и др. Из алкалоидов растительного происхождения наиболее известны — налоксон, пентазоцин и налтрексон. Два последних обладают свойствами частичных антагонистов. Эндогенные пептиды — эндорфины, энкефалины и динорфины —взаимодействуют аналогично морфину и относятся к агонистам ОР.
ОР гетерогенны по составу, и классифицируют их на основе взаимодействия со специфическими лигандами. Они подразделяются на субтипы: мю-, дельта-, каппа-, сигма- и эпсилон-рецепторы. Принято считать, что морфин выявляет мю-тип рецепторов, Ала, Лей-энкефалин, дельторфины и некоторые другие аналоги энкефалинов — дельта-рецепторы, кетоциклазоцин и динорфины —каппа-рецепторы, N-аллилнорияклазоцин — сигма-рецепторы, а р-эндорфин —мю- и эпсилон-рецепторы. Вместе с тем следует помнить, что эти лиганды не обладают абсолютной специфичностью и могут частично взаимодействовать с разными субтипами О Р.
Связывание радиолигандов свидетельствует о различном распределении субтипов ОР по структурам головного мозга.
Для изучения биохимических характеристик опиатных рецепторов многие годы предпринимались попытки выделить их в нативном состоянии из биологических мембран мозга. Однако ОР оказались трудным объектом, они инактивировались под действием ионных детергентов, которые ранее успешно применялись для солюбилизации других рецепторов. Кроме того, попытка полного удаления липидного окружения из препаратов ОР также вела к инактивации.
Лишь использование неионных детергентов типа дигитонинаглиоксихолата и CHAPS — позволяет солюбилизировать из мембран белки рецепторного комплекса с сохранением 50-70% опиатсвязывающей активности, ОР-рецепторы оказались ли-попротеидами. Роль липидных компонентов и окружения ОР в связывании опиоидов пока не ясна, однако, присутствие сульфоцереброзидов существенно увеличивает уровень связывания агонистов с рецепторами, и было сделано предположение о том, что сульфоцереброзиды входят в активный центр ОР.
Собственно рецепторные лиганд-связывающие компоненты ОР имеют молекулярную массу порядка 45000-66000 дальтон. В целом ОР подобны другим метаботропным рецепторам. Взаимодействие с лигандом ведет к включению G-белков и, в конечном счеге, к подавлению аденилатциклазы. Интересно, что молекула рецептора обладает дефосфорилирующей активностью. Она усиливается теми же факторами, которые усиливают связывание опиатов. При этом дефосфорилирование изменяет ли-ганд-рецепторное взаимодействие.
Известно также, что активация опиатных рецепторов сопровождается повышением К+ проводимости мембраны и/или снижением Са+ проводимости мембраны, что ведет к принципиально общему конечному результату — уменьшению входа Са+ в пресинаптические окончания во время прохождения потенциала действия и к снижению количества высвобождаемого нейромедиатора или гиперполяризации постсинаптической мембраны.
Ферменты-рецепторы. Название этого типа рецепторов вызывает некоторое недоумение, ибо конечный компонент мета-ботропных рецепторов является, как правило, ферментом, например циклазой или фосфолипазой. Однако последние служат лишь компонентом комплекса белков, составляющих метаботропный рецептор. Медиатор при этом не взаимодействует непосредственно с ферментом. Существует, однако, ряд систем межнейрональной передачи сигнала, когда медиатор прямо действует на фермент. Приведем лишь один пример. Относительно новый нейромедиатор нитроксид прямо активирует гуанилатциклазу и синтез цГМФ.
Выводы
1. Обязательным звеном передачи нервного импульса в химических синапсах являются рецепторы — образования, состоящие из белков и гликолипидных компонентов, которые с высокой специфичностью связывают нейромедиатор, меняют конформацию и обеспечивают трансформацию сигнала в изменения ионных потоков через мембрану и в образование вторичных мессенджеров в клетке.
2. По типу вызываемых медиатором процессов рецепторы делятся на две категории: 1) быстродействующие, содержащие в своей структуре ионный канал, открытие которого ведет к изменению потенциала мембраны; 2) медленнодействующие, состоящие из компонентов, периодически связывающихся друг с другом, которые после взаимодействия с нейромедиатором запускают цепь реакций, образующих вторичные молекулы — посредники, циклические нуклеотиды, диацилглицерол, ино-зитолфосфаты и др.
3. Одни и те же медиаторы в разных синапсах могут взаимодействовать с рецепторами разных типов и разных подтипов.
4. Современный уровень понимания структуры и механизма действия многих рецепторов таков, что оказалось возможным "собирать" активные рецепторы из компонентов вне клеток — в мембранах липосом — и даже индуцировать с помощью соответствующих мРНК их синтез, встраивание в мембраны и действие в клетках, не содержащих ранее этих образований.
5. Действие ряда важнейших фармакологических агентов, используемых при лечении заболеваний ЦНС, направлено на рецепторы нейромедиаторов — их активацию или подавление.
Похожие работы

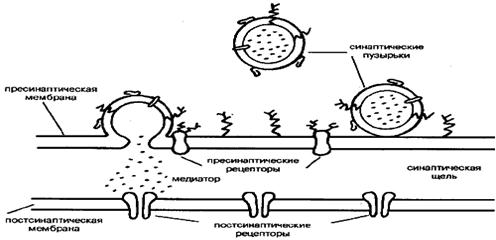

... , накапливающие и хранящие медиатор вплоть до момента его освобождения в синаптическую щель. Мембрана синаптических пузырьков содержит разнообразные белки, предположительно участвующие в освобождении медиатора. 3. Материальная основа высшей нервной деятельности. Нервная система - совокупность структур в организме животных и человека, объединяющая деятельность всех органов и систем и ...
... в состоянии продолжать жить, несмотря на указания совести о неправильности жизни…». На этом тезисе мы закончили предыдущую главу книги. Не развивая его, поскольку это не входит в достаточно узкую заявленную тематику оружия геноцида, обратимся к внутренним механизмам, которые включаются у людей после принятия психотропов. Мы знаем от чего люди убегают (от своей совести). Но за чем же люди гонятся? ...

... материалы хорошо описываются в рамках квантово-механической фононной Модели строения и функционирования клеточных мембран, что позволяет утверждать: “ФОНОН – КВАНТ биологической (клеточной) мембраны”. Модель пригодна для объяснения широкого круга наблюдаемых явлений. При этом наблюдаемые явления описываются в рамках единого понятийного аппарата и не требуют специфических допущений для описания ...
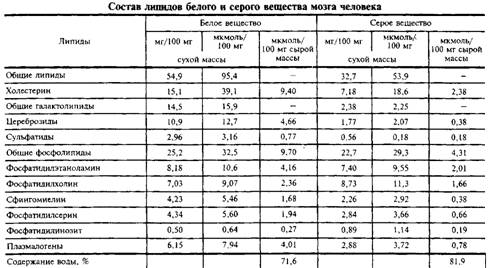
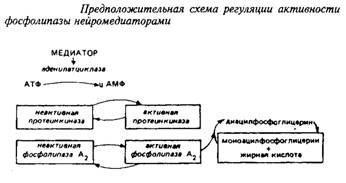

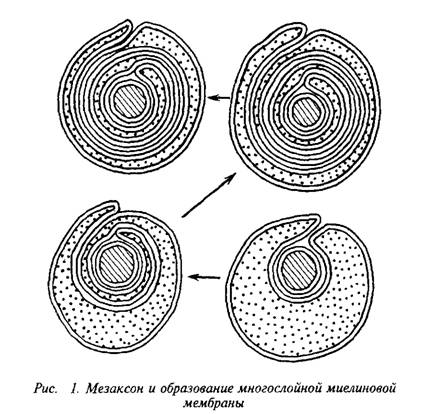
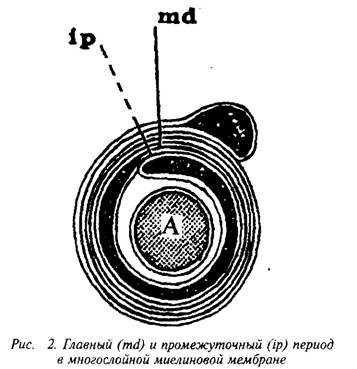
... и инозитолтрифосфат подвергаются химическим превращениям, требующим АТФ и ЦТФ и приводящим к восстановлению три-фосфоинозитида. Таким образом, цикл замыкается и уровень полифосфоинозитидов в мембране восстанавливается. 7. МИЕЛИН В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ Мозг человека содержит 120 г миелина, что составляет одну треть его сухой массы. Миелин – уникальное образование, организация которого ...
0 комментариев