Навигация
Действие ренина, ангиотензина и кининов на почки
21779
знаков
0
таблиц
0
изображений
Министерство образования Российской Федерации
Пензенский Государственный Университет
Медицинский Институт
Кафедра Терапии
РЕФЕРАТ
на тему:
«Действие ренина, ангиотензина и кининов на почки»
Пенза
2010
План
1. Действие ренина и ангиотензина
2. Действие кининов
Литература
1. Действие ренина и ангиотензина
Известно, что в гранулированных клетках юкстагломерулярного аппарата (ЮГА) главным образом корковых нефронов вырабатывается ренин — протеолитический фермент, расщепляющий один из а2-глобулинов (ангиотензиноген) с образованием декапептида ангиотензина I. Последний мало активен, но под влиянием так называемого превращающего фермента он переходит (главным образом в легких, но отчасти в почках) в гораздо более активный октапептид ангиотензин П. Этим объясняется слабый эффект ангиотензина I при введении его в почечную артерию по сравнению с ангиотензином II в отношении почечного кровотока, но одинаковый эффект при внутривенном введении, когда ангиотензин I до попадания в почку проходит через малый круг кровообращения.
Интересно, что в различных тканях животных и человека (матка, плацента, надпочечники, подчелюстная железа) вырабатываются изоферменты (изозимы) ренина, имеющие тот же субстрат. Под влиянием изоферментов ренина, полученных из почки и матки собаки, почки крысы и подчелюстной железы мыши, в крови образуется один и тот же ангиотензин I, что подтверждено иммунологически.
Мы не будем анализировать огромную литературу о ЮГА и факторах, регулирующих секрецию ренина и образование ангиотензина. Этим вопросам посвящен ряд обзоров (Серебровская Ю. А., 1972, 1974; УшкаловА. Ф., 1972; Ратнер М. Я. и др., 1973). Главными факторами, влияющими на секрецию ренина, являются нарушения гемодинамики с изменением почечного кровотока, содержание натрия в крови и моче дистального отдела канальцев, уровень секреции минералокортикоидов (обратная связь). Ведущую роль при этом играют клетки плотного пятна (macula densa), обладающие баро- и хемоцептивной функцией. Что касается значения нервных (симпатических) влияний на секрецию ренина, то по этому вопросу нет единого мнения. Изучая роль почек в развитии экспериментальной гипертонии, В. Н. Черниговский и его сотрудники установили, что нервные влияния стимулируют образование ренина, а денервация почек снижает его образование независимо от состояния почечного кровообращения (Черниговский В. Н., 1955; Ратнер М. Я., и др., 1957). Это нашло подтверждение в ряде недавних работ, показавших, что у собак с денервированной почкой содержание ренина оказывается сниженным. Однако усиленная секреция ренина в ответ на ограничение соли в диете наблюдалась на фоне введения ганглиоблокаторов и адренолитиков, а также после денервации почки.
Повышенный уровень ангиотензина в крови по механизму обратной связи снижает секрецию ренина. Это было показано на собаках, а также на овцах, которым ангиотензин вводили как внутривенно, так и в почечную артерию. Значение торможения секреции ренина ангиотензином для поддержания постоянства концентрации последнего в крови продемонстрировано недавно на математической модели ренин-ангиотензиновой системы (Колчанов Н. А., Поляк М. Г., 1974). Секреция ренина у собак снижается также при введении АДГ в дозах, не влияющих на гемодинамику. В связи с этим представляют интерес сообщения о влиянии ангиотензина на секрецию АДГ. Согласно наблюдениям некоторых авторов, введение собакам ренина или ангиотензина повышало содержание АДГ в крови. Этот эффект имел место при внутривенном введении ангиотензина, но был особенно выражен при его введении в сонную артерию. Найдено повышение концентрации АДГ в крови при инфузии ангиотензина в латеральный желудочек мозга у крыс. На основании подобных наблюдений высказывалось предположение об участии АДГ в проявлении действия ангиотензина. Однако другие исследователи, сравнивая концентрирующую функцию почек или введении ангйотензина внутривенно и в сонную артерию, пришли к выводу об отсутствии прямого влияния ангиотензина на секрецию АДГ или на его периферическое действие. Не было подтверждено повышение содержания АДГ в плазме при внутривенном введении ангиотензина собакам. В связи с этим вопрос о взаимоотношениях между ангиотензином и АДГ остается дискуссионным.
Из трех основных направлений, по которым действует ангиотензин: сосудистая система, надпочечники, почки,— остановимся только на последнем. Правда, как правильно отметили в своем обстоятельном обзоре А. М. Вихерт и А. Ф. Ушкалов (1971), влияние ангиотензина на функцию почек является наиболее трудным для понимания. Экспериментальные данные по этому вопросу и выводы весьма разнообразны, что связано с неодинаковым действием ангиотензина в зависимости от дозы, исходного состояния и вида животного на различные сосудистые участки в почках и канальцевые процессы (Бранчевский Л. Л., 1972).
У интактных животных и человека физиологические дозы ангиотензина снижают, как правило, диурез и натрийурез (Колпаков М. Г. и др., 1974), что является результатом уменьшения почечного кровотока и клубочковой фильтрации в связи с повышением тонуса афферентных артериол, а впоследствии и усиления реабсорбции натрия из-за стимуляции альдостерона. Сходный эффект наблюдался в опытах с изолированной почкой кролика, когда добавление ангиотензина к перфузируемой жидкости оказывало тонизирующее влияние на эфферентные артериолы. Не исключается и прямое стимулирующее действие ангиотензина на канальцевый транспорт натрия, показанное на срезах коркового вещества почек крыс, причем это влияние сохранялось на фоне угнетения Nа, К-АТФазы строфантином, а потому оно, возможно, связано с иным натриевым насосом.
В лаборатории, руководимой М. Г. Колпаковым, было получено резкое снижение натрийуреза после внутривенного введения собакам ангиотензина в условиях хронического опыта. При этом снижались почечный кровоток и клубочковая фильтрация, но в большей степени усилилась реабсорбция натрия. Концентраций альдостерона в плазме не изменилась (Колпаков М. Г. и др., 1975). Авторы считают, что имеется прямое влияние ангиотензина на почки, которое может реализоваться с участием физических факторов, например изменения гидростатического давления в перитубулярных капиллярах, либо путем усиления активного транспорта натрия, или же благодаря перераспределению почечного кровотока к юкстамедуллярной зоне, нефроны которой в большей степени реабсорбируют натрий.
С другой стороны, если исходная фильтрация снижена вследствие падения системного давления или сужения почечной артерии, ангиотензин ведет к сужению эфферентных артериол, в результате чего повышается фильтрационная фракция и создаются условия для усиления фильтрации. Не исключено, что такое регулирующее влияние ангиотензина на фильтрацию, а следовательно, и выделение натрия, оказывается возможным благодаря тому, что образуемый в ЮГА ренин может транспортироваться с помощью специального переносчика в цитоплазму клеток афферентных артериол, где и образуется небольшое количество ангиотензина. Если фильтрация усилена, загрузка дистального отдела повышена, и натрий не успевает реабсорбироваться в достаточном количестве в восходящем отделе петли Генле. Тогда увеличение его концентрации в начальной части дистального канальца активирует образование переносчика и транспорт ренина в афферентные артериолы, что ведет к повышению их тонуса и ограничению фильтрации. При снижении экскреции натрия его концентрация в дистальном канальце падает, ренин в меньшей степени транспортируется в цитоплазму афферентных артериол, их тонус снижается, и усиливается фильтрация. В то же время накапливаемый в ЮГА ренин реализуется через клубочковый кровоток и действует на эфферентные сосуды. Участие ангиотензина в поддержании клубочково-канальцевого баланса заключается, по мнению Leyssac (1967), не только в изменении фильтрации, но и в уменьшении скорости проксимальной реабсорбции натрия. Однако последнее впоследствии не подтвердилось.
Помимо влияния на клубочковый кровоток, ангиотензин, по-видимому, принимает участие в распределении внутрипочечного кровотока и, возможно, является его физиологическим регулятором. При перфузии изолированной почки собаки было найдено, что вначале кровоток наружного коркового слоя составлял 79%, а внутреннего—21% от общего эффективного кровотока. Постепенно кровоток смещался в сторону внутреннего, который на 150-й минуте инфузии составил уже 34%. Оказалось, что к этому времени в почке исчезал субстрат ренина — ангиотензиноген. При его добавлении в питающую жидкость восстанавливалось прежнее распределение кровотока. Интересно, что ангиотензин II, введенный извне, не дает такого эффекта. Это говорит в пользу того, что эндогенный ангиотензин является внутрипочечным гормоном, участвующим в распределении почечного кровотока. Циркулирующий в крови ангиотензин такой функцией не обладает. Изложенные данные были подтверждены в более физиологических условиях на собаках, у которых внутрипочечное распределение кровотока между корковым и мозговым веществом изучали методом вымывания радиоактивного ксенона. При сужении почечной артерии у интактных собак кровоток перераспределялся так, что меньше всего страдал корковый кровоток. Если же предварительно снижали образование ренина (введение внутрь больших количеств соли и ДОКСА), то в ответ на сужение почечной артерии корковый кровоток уменьшался пропорционально общему и даже, наоборот, имелась тенденция к перераспределению крови в пользу мозгового кровотока. Авторы признают существование двух механизмов ауторегуляции внутрипочечного кровотока: один, ренин-ангиотензиновый, стоит на страже наружного кортикального кровотока, другой, не связанный с ренином, обеспечивает юкстамедуллярный кровоток. По мнению некоторых авторов, ренин-ангиотензиновая система не играет существенной роли в ауторегуляции почечной гемодинамики.
При введении ангиотензина животным в дозах, превышающих физиологические, а также при действии малых доз у больных с гипертонией или у животных с предварительной нагрузкой солью наблюдается натрий-уретйческое и диуретическое действие (Бранчевский Л. Л., 1973). Этот эффект может, вероятно, в некоторых случаях зависеть от уменьшения капиллярного кровотока в корковом веществе, что снижает проксимальную реабсорбцию (Федотов А. М., Федоров В. И., 1974). Однако главным механизмом натрийуретического и диуретического действия ангиотензина является прямое угнетающее влияние на канальцевый транспорт натрия. Это было показано на срезах коркового вещества почек крыс и кроликов, что исключало влияние через изменение фильтрации или перитубулярного кровотока. Кроме того, получено одностороннее диуретическое и натрийуретическое действие ангиотензина при его введении в портальный кровоток курицы и жабы. При этой методике, как известно, вещество не попадает в клубочки, и наблюдаемые эффекты обязаны только канальцевому действию. В опытах на собаках с введением ангиотензина в почечную артерию и применением метода stop flow было установлено прямое угнетающее влияние на реабсорбцию натрия в дистальном, но не в проксимальном отделе канальцев. При перфузии через изолированные проксимальные отделы канальцев кроликов ангиотензин не влиял на реабсорбцию жидкости. Прямое действие ангиотензина при введении в кровоток одной из почек было подтверждено и на кроликах. Затем с помощью микропункции канальцев у крыс удалось наблюдать снижение реабсорбции натрия в дистальном отделе, если ангиотензин вводили в перитубулярный кровоток. При введении же даже большей концентрации в просвет канальца эффекта не было. Следовательно, ангиотензин действует на дистальный отдел канальца со стороны базальной поверхности клеток. Вместе с тем имеются указания на то, что натрийуретическое действие ангиотензина обязано его влиянию на проксимальный отдел канальца и восходящее колено петли Генле. В опытах с одновременной микроперфузией просвета и перитубулярных капилляров проксимальных отделов канальцев ангиотензин II, вводимый в перитубулярную жидкость, вызывал двухфазный эффект: повышение реабсорбции натрия при низких концентрациях и угнетение реабсорбции —при более высоких. При введении в просвет канальца действие было менее выраженным. Следовательно, ангиотензин II оказывает прямое влияние на проксимальные отделы канальцев и главным образом со стороны перитубулярной поверхности клеток.
Определенные сведения о непосредственном влиянии ангиотензина на канальцевые процессы могли быть получены в модельных опытах. Однако согласно одним авторам, ангиотензин не влияет на транспорт электролитов через стенку мочевого пузыря, кожи или кишки амфибий, тогда как другие наблюдали изменения транспорта, характер которых зависел от используемых концентраций. В связи с этим вряд ли эти данные существенно дополняют обсуждаемый вопрос.
При длительном введении ангиотензина повышенная в первые дни экскреция натрия и воды постепенно снижается, и наступает задержка натрия. Так, согласно исследованию И. Ш. Штеренталя (1968, 1969), на 4— 5-е сутки введения ангиотензина внутрь диурез после первоначального усиления вернулся к исходным величинам, а натрийурез снизился. Автор объясняет это превалированием натрийзадерживающего действия альдостерона, секреция которого постепенно стимулировалась ангиотензнном. В этих опытах наблюдалось и характерное для альдостерона перераспределение натрия в организме с увеличением его депонирования. Количество обменоспособного натрия не изменилось.
Похожие работы
... их в крови (сульфаты). Аминокислоты. Реабсорбция аминокислот происходит также по механизму сопряженного с Na+ транспорта. Профильтровавшиеся в клубочках аминокислоты на 90% реабсорбируются клетками проксимального канальца почки. Этот процесс осуществляется с помощью вторично-активного транспорта, т.е. энергия идет на работу натриевого насоса. Выделяют не менее 4 транспортных систем для переноса ...
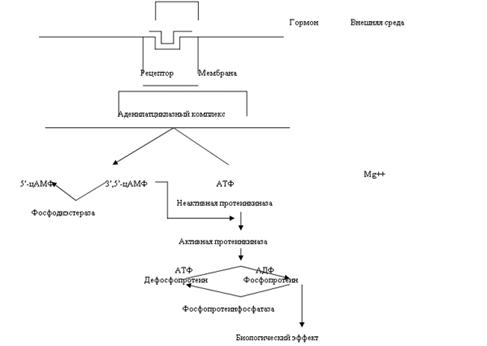

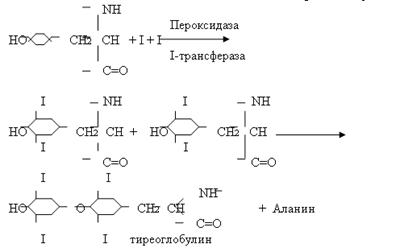

... эффект опосредуется связыванием ГР-РФ с рецепторами плазматической мембраны соматотрофов и активацией трех систем вторичных посредников. С их помощью происходит усиленный синтез мРНК СТГ. Этот эффект действия водорастворимого гормона на ядерный аппарат клетки объясняют фосфорилированием и дефосфорилированием определенной группы протеинкиназ, которые действуют на генетический аппарат клетки. На ...
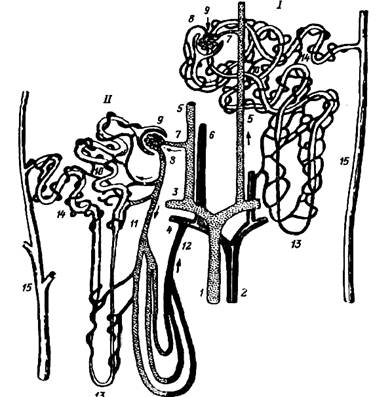
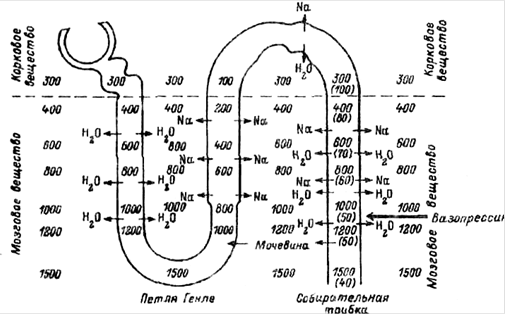
... состояния водно-солевого баланса в организме. Следствием центрального анализа является изменение питьевого и пищевого поведения, перестройка работы желудочно-кишечного тракта и системы выделения (прежде всего функции почек), реализуемая через эфферентные звенья регуляции. Последние представлены нервными и, в большей мере, гормональными влияниями. Метаболическая функция почек. Метаболическая ...
... а также других эндокринных желез, кроме гипофиза и надпочечников. Наряду с этим многие фармакологические вещества оказывают прямое влияние на почечные механизмы мочеобразования. Исследование непосредственного воздействия на почки до последних лет проводилось главным образом в отношении веществ, для которых оно является основным (мочегонные средства, АДГ, альдостерон). В отношении большинства ...
0 комментариев