Наиболее изученными из ионотропных рецепторов глутамат являются NMDA-рецепторы. Им отводится особая роль в регуляции нейрональной возбудимости, синаптической пластичности (Cotman C.W. еt al., 1987), а также в патогенезе эпилепсии и судорог (Chapman A.G. 1998; Bradford H.F. 1995; Dingledine R. et al., 1990).
NMDA-рецепторы представляют собой тетрамерный комплекс, формируемый комбинацией двух субъединиц: NR1 и NR2 (Laube B. et al., 1998). Каждая из субъединиц NMDA-рецептора представлена рядом изоформ, возникающих в результате альтеранивного сплайсинга. Различные их комбинации формируют рецепторы, различающиеся по фармакологическому профилю, чувствительности к Mg2+ и свойствам каналов (Sucher N.J. et al., 1996).
Ионный канал, образованный субъединицами, высоко проницаем для K+, Na+, Ca2+ (McBain C.J., Mayer M.L., 1994) и заблокирован ионом Mg2+ потенциалзависимым способом (Nowak L. et al., 1984). Деполяризация постсинаптической мембраны (от -50 до -30 мВ), вызванная активацией других глутаматных ионотропных рецепторов, устраняет «магниевый блок» и приводит к открытию канала. Ионофор NMDA-рецепторов является участком связывания так называемых «канальных» блокаторов (фенциклидина, кетамина, мемантина, МК-801) (Dingledine R. et al., 1999; McBain C.J., Mayer M.L., 1994).
Применение радиолигандного анализа показало наибольшую плотность NMDA-рецепторов в конечном мозге, прежде всего в гиппокампе, коре больших полушарий, миндалине и стриатуме (Maragos W.F. et al., 1988; Monaghan D.T., Cotman C.W., 1985). Следует отметить, что именно эти структуры прежде всего ответственны за память и обучение в традиционном понимании этих слов и ассоциированы с сенсорной функцией, осуществление которой требует повышенного ресурса синаптической пластичности (Беспалов А.Ю., Звартау Э.Э., 2000). Представляет также интерес то, что указанные структуры обладают низким порогом эпилептизации и высокой степенью возбудимости.
Во всех слоях гиппокампа выявлен высокий уровень NMDA-рецепторов, за исключением тел нейронов пирамидного и гранулярного слоев, а также striatum lucidum (терминальная зона мшистых волокон гиппокампа). Среди кортикальных областей ассоциативные зоны коры часто имеют большую плотность рецепторов, чем проекционные зоны. Фронтальная, инсулярная, пириформная, периринальная и передняя поясная кора также содержат больше количество рецепторов в отличие от височной, затылочной, париетальной и задней поясной участков коры. Гранулярные корковые области имеют выраженную ламинарность распределения NMDA-рецепторов. Так, во внешних слоях I-III и слое Vа париетальной коры показана бoльшая плотность рецепторов, чем в других корковых слоях.
NMDA-рецепторы опосредуют возбуждающее действие глутамата, однако, оно не распространяется на все виды активации нейрона. Они не участвуют в возникновении быстрого и кратковременного возбуждения, с которым связаны физиологические реакции (например, защитный рефлекс одергивания и др.). Быстрые возбуждающие постсинаптические потенциалы, характерные для подобных реакций, формируются при участии ионотропных не-NMDA-рецепторов. Роль же NMDA-рецепторов велика в патологии нейронов, их гиперактивации и эпилептизации, так как обеспечивают усиленную и длительную активацию нейронов. Показано, что селективный антагонист NMDA-рецепторов – амино-5-фосфовалериановая кислота – предупреждает и ослабляет гиперактивацию нейронов и судорожные реакции, но существенно не влияет на физиологические двигательные реакции (Крыжановский Г.Н., 1997).
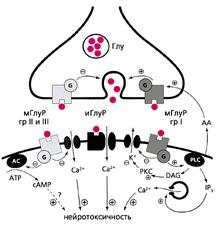
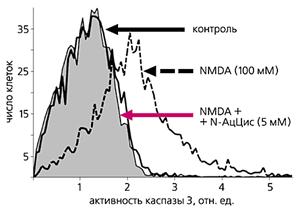
В физиологических условиях NMDA-рецепторы активируются миллимолярными концентрациями глутамата, который присутствует в синаптической щели в течение нескольких миллисекунд (Clements J.D. et al., 1992). При патологической импульсации рецепторы активируются микромолярными концентрациями, но в течение значительно большего времени (Globus M.Y. et al., 1988). В результате этого происходит увеличение концентрации Са2+ в клетках и накопление ионов К+ во внеклеточном пространстве. "Кальциевая перегрузка" нейронов и активация Са2+-зависимых процессов (повышение активности протеаз, киназ, эндонуклеаз, липооксигеназ, фосфолипазы А2 и др. ферментов) ведет к значительным изменениям в метаболизме и генетическом аппарате клетки, неконтролируемому действию свободных радикалов и может привести к необратимой клеточной гибели (Choi D.W., 1985). В настоящее время считается, что поступление ионов Са2+ внутрь клетки через каналы NMDA-рецепторов является ключевым событием в реализации токсических эффектов глутамата (Bading H. et al., 1993). Так, установлено, что эквивалентный подъем внутриклеточной концентрации Са2+, поступающего через потенциал-зависимые кальциевые каналы, приводит к меньшим нейрональным повреждениям (Dubinsky J.M. et al. 1991). Насыщение нейронов ионами кальция служит триггером для мобилизации ионов из внутриклеточного депо, что также имеет значение в токсических эффектах глутамата (Frandsen A. et al., 1992; Lei S.Z. et al., 1992). Повышение концентрации внеклеточного К+ рассматривается многими авторами как один из важнейших механизмов вовлечения нейронов в эпилептический процесс (Dichter M.A. et al., 1972; Hillman H., 1970). Орошение срезов мозга средой, обогащенной ионами К+, как известно, является моделью эпилептогенеза in vitro. Так, в поле СА1 гиппокампа повышение К+ индуцирует спонтанные электрофизиологические судороги с тонической и клонической фазами, характерными для фокальной эпилепсии (Traynelis S.F., Dingledine R., 1988).
Дополнительным результатом активации NMDA-рецепторов является внутриклеточная продукция активных форм кислорода, прежде всего супероксид-аниона и гидроксид-радикала. При дефиците аргинина, субстрата нитроксидсинтазы, также может образовываться супероксид-анион. В условиях избыточного образования различных радикалов возможно взаимодействие вторичного мессенджера – оксида азота и супероксида с образованием пероксинитрита, обладающего исключительно высоким окислительным потенциалом (Ha H.C., Snyder S.H., 2000).
Таким образом, гиперактивация NMDA-рецепторов, несомненно, вносит определенный вклад в судорожную активность и связанную с ней гибель нейронов, что также подтверждается большим количеством экспериментальных данных о выраженном противосудорожном эффекте антагонистов NMDA-рецепторов, проявляющемся на различных моделях судорог. Так, существуют указания на усиление антагонистами NMDA-рецепторов лечебного эффекта бензодиазепинов и барбитуратов при данных состояниях (Федонюк В.П. и соавт., 2001).
Глутамат играет важную роль в процессах дифференцировки, миграции и жизнеспособности нейронов, в основном, через усиление входящего тока Ca++ (Hack and Balazs 1994 , Yano et al. 1998 ).
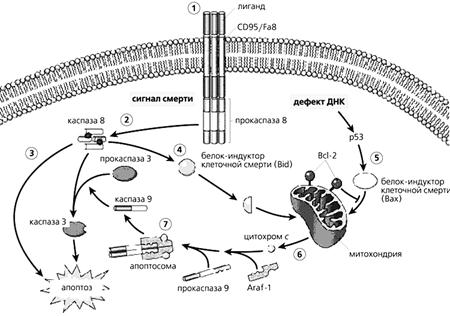
Блокада НМДА рецепторов в пренатальный период [дизоцилпином (MK-801), фенциклидином, этанолом] может вызыветь апоптоз в уязвимых нейронах (селективная уязвимость, зависящая от стадии развития) (Ikonomidou et al. 1999 ).
Перевод: Иванов
Похожие работы

... роль нейрорецепторов сводится к созданию специфических информационных входов, организующих единый функциональный ансамбль нейронов. Именно совокупность рецепторов определяет лицо клетки и ее реакции на поступление разнообразных химических сигналов. Молекулярные механизмы, лежащие в основе модуляции эффективности синаптической передачи, в которых важную роль играют рецепторные процессы, имеют ...
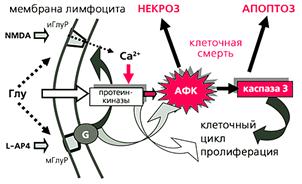
... многих стадиях клеточного цикла и в процессах пролиферации [4]. Особенно важны эти реакции для клеток иммунной системы. Значит, в ряде случаев активность каспазы 3 не обязательно означает начало апоптоза, а может быть связана с пролиферацией лимфоцитов. Глутаматные рецепторы иммуннокомпетентных клеток История открытия и изучения глутаматных рецепторов накопила массу примеров их причастности к ...

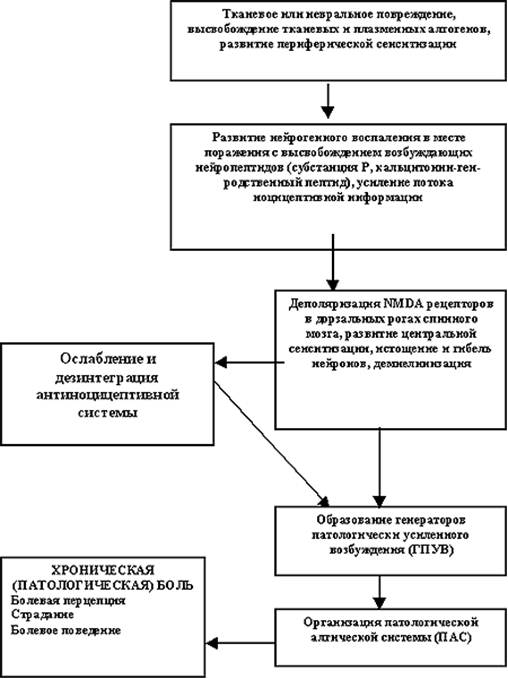
... . 6. Формирование патологической алгической системы (ПАС), включающей различные уровни центральной нервной системы и определяющей течение и характер всех компонентов патологической боли: болевой перцепции, страдания и болевого поведения. Хроническая боль как общепатологическая проблема Что же представляет собой хроническая (патологическая) боль с позиций общей патологии? В учении о болезни ...
... лигандами, большая часть массы белка канала находится на цитоплазматической стороне мембраны. Наиболее вероятной формой объединения субъединиц является тетрамер. Разнообразие субъединиц Характерной особенностью ионных каналов является широкое разнообразие изоформ субъединиц. Существует больше дюжины вариантов субъединиц никотинового АХР и еше большее число субъединиц калиевого канала и ...
0 комментариев