Навигация
Окислительные пути распада углеводов в тканях
57980
знаков
0
таблиц
0
изображений
2.1. Окислительные пути распада углеводов в тканях
Важнейшими функциями моносахаридов в организме являются энергетическая и пластическая; обе эти функции реализуются в ходе окислительного распада моносахаридов в клетках. При окислении углеводов выделяется 4,1 ккал/г ( около 17 кДж/г ) свободной энергии и за счет окисления углеводов человек покрывает 55-60% своих общих энергозатрат. В ходе окисления углеводов образуется большое количество промежуточных продуктов распада, которые используются для синтеза различных липидов, заменимых аминокислот и др. необходимых клеткам соединений. Кроме того, при окислении углеводов в клетках идет генерация восстановительных потенциалов, которые в дальнейшем используются ими в восстановительных реакциях биосинтезов, в процессах детоксикации, для контроля уровня перекисного окисления липидов и др.
Главным моносахаридом, подвергающимся окислительным превращениям в клетках, является глюкоза, поскольку именно она в наибольших количествах поступает из кишечника во внутреннюю среду организма, именно она синтезируется при глюконеогенезе или образуется в свободном виде или же в виде фосфорных эфиров при расщеплении гликогена. Роль других моносахаридов менее значительна, так как их количество, поступающее в клетки в количественном отношении сильно варьирует в зависимости от состава пищи.
Известно несколько метаболических путей окисления глюкозы, главными из которых являются:
а) аэробное расщепление до углекислого газа и воды;
б) анаэробное окисление до лактата;
в) пентозный путь окисления;
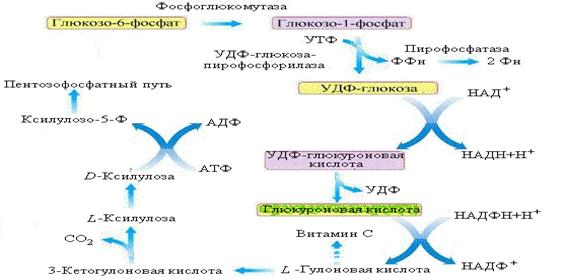
г) окисление с образованием глюкуроновой кислоты.
Глубина окислительного расщепления молекулы глюкозы может быть различной: от окисления одной из концевых группировок молекул до карбоксильной группы, что происходит при образовании глюкуроновой кислоты, до полной деградации молекулы глюкозы при ее аэробном распаде.
2.1.1. Аэробное окисление глюкозы
В клетках аэробных организмов основным, по крайней мере в отношении общего количества расщепляющейся глюкозы, является ее аэробный распад до углекислого газа и воды. При расщеплении 1 М глюкозы ( 180 г ) в аэробных условиях выделяется 686 ккал свободной энергии. Сам процесс аэробного окисления глюкозы можно разделить на 3 этапа:
1. Расщепление глюкозы до пирувата.
2. Окислительное декарбоксилирование пирувата до ацетил-КоА.
3. Окисление ацетила в цикле Кребса ( ЦТК ), сопряженное с работой цепи дыхательных ферментов.
2.1.1.1. Расщепление глюкозы до пирувата
По современным представлениям первый этап окисления глюкозы протекает в цитозоле и катализируется надмолекулярным белковым комплексом - гликолитическим метаболоном, включающим в себя до десятка отдельных ферментов.
Первый этап окисления глюкозы может быть в свою очередь разделен на 2 стадии. В реакциях первой стадии происходит фосфорилирование глюкозы, изомеризация остатка глюкозы в остаток фруктозы, дополнительное фосфорилирование уже фруктозного остатка и, наконец. расщепление гексозного остатка на два остатка фосфотриоз:
Эта реакция катализируется ферментом гексокиназой. В качестве фосорилирующего агента в клетке используется АТФ. Реакция сопровождается потерей свободной энергии порядка 5,0 ккал/моль и в условиях клетки является необратимой.
Регуляция работы первого этапа аэробного расщепления глюкозы осуществляется с помощью термодинамических механизмов и с помощью механизмов аллостерической модуляции регуляторных ферментов, принимающих участие в работе этого метаболического пути.
С помощью термодинамических механизмов осуществляется контроль направления потока метаболитов по данному метаболическому пути. В описанную систему реакций включены три реакции, в ходе которых теряется большое количество энергии: гексокиназная ( G0= - 5,0 ккал/моль ), фосфофруктокиназная ( G0= -3,4 ккал/моль ) и пируваткиназная ( G0= - 7,5 ккал/моль ). Эти реакции в клетке практически не обратимы, в особенности пируваткиназная реакция, и за счет их необратимости процесс становится необратимым в целом.
Интенсивность потока метаболитов по рассматриваемому метаболическому пути контролируется в клетке за счет изменения активности включенных в систему аллостерических ферментов: гексокиназы, фосфофруктокиназы и пируваткиназы. Таким образом, пункты термодинамического контроля метаболического пути одновременно являются и участками, на которых осуществляется регуляция интенсивности потока метаболитов.
Главным регуляторным звеном системы является фосфофруктокиназа. Активность этого фермента подавляется высокими концентрациями АТФ в клетке, степень аллостерического ингибирования фермента АТФ усиливается при высоких концентрациях цитрата в клетке. АМФ является аллостерическим активатором фосфофруктокиназы.
Гексокиназа ингибируется по аллостерическому механизму высокими концентрациями Гл-6-ф. В этом случае мы имеем делом с работой сопряженного регуляторного механизма. В клетке после угнетения активности фосфофруктокиназы высокими концентрациями АТФ накапливается Фр-6-ф, а значит накапливается и Гл-6-ф, поскольку реакция, катализируемая фосфогексоизомеразой, легко обратима. В таком случае повышение концентрации АТФ в клетке ингибирует активность не только фосфофруктокиназы, но и гексокиназы.
Очень сложно выглядит регуляция активности третьей киназы пируваткиназы. Активность фермента стимулируется Гл-6-ф, Фр-1,6-бф и ФГА по аллостерическому механизму - так называя активация предшественником. В свою очередь, высокие внутриклеточные концентрации АТФ,НАДН,цитрата, сукцинил-КоА и жирных кислот угнетают активность фермента по аллостерическому механизму.
В целом, расщепление глюкозы до пирувата тормозится на уровне 3 указанных киназ при высокой концентрации АТФ в клетке,т.е. в условиях хорошей обеспеченности клетки энергией. При недостатке энергии в клетке активация расщепления глюкозы достигается,во первых, за счет снятия аллостерического ингибирования киназ высокими концентрациями АТФ и аллостерической активации фосфофруктокиназы АМФ и, во-вторых, за счет аллостерической активации пируваткиназы предшественниками: Гл-6-Ф, Фр-1,6-бф и ФГА.
Каков смысл ингибирования цитратом фосфофруктокиназы и цитратом и сукцинил-КоА - пируваткиназы? Дело в том, что из одной молекулы глюкозы образуется две молекулы ацетил-КоА, который затем окисляется в цикле Кребса. Если в клетке накапливаются цитрат и сукцинил-КоА, значит цикл Кребса не справляется с окислением уже наработанного ацетил-КоА и есть смысл притормозить его дополнительное образование, что и достигается ингибированием фосфофруктокиназы и пируваткиназы.
Наконец, угнетение окисления глюкозы на уровне пируваткиназы при повышении концентрации жирных кислот направлено на сбережение глюкозы в клетке в условиях, когда клетка обеспечена другим, более эффективным видом энергетического топлива.
Похожие работы
... механизмах синтеза гликогена, во всех окислительных путях превращения глюкозы и в синтезе других моносахаридов,необходимых для клетки. Место, которое занимает данная реакции в обмене глюкозы позволяет ее счиатать ключевой реакцией обмена углеводов. Гексокиназная реакция необратима (G= -16,7 кДж/моль), поэтому для превращения глюкозо-6-фосфата в свободную глюкозу в клетках печени и почек ...

... α,d – глюкоза глюкозо – 6 – фосфат С образованием глюкозо – 6 – фосфата пути гликолиза и гликогенолиза совпадают. Глюкозо – 6 – фосфат занимает ключевое место в обмене углеводов. Он вступает в следующие метаболические пути: глюкозо – 6 – фосфат глюкоза + Н3РО4 фруктозо – 6 – фосфат пентозный путь распада (поступает в кровь и др. ...
... . Динамика химических превращений, происходящих в клетках, изучается биологической химией. Задачей физиологии является определение общих затрат веществ и энергии организмом и того, как они должны восполняться с помощью полноценного питания. Энергетический обмен служит показателем общего состояния и физиологической активности организма. Единица измерения энергии, обычно применяемая в биологии и ...
... затратах энергии; 2) при переменных затратах энергии и 3) при затратах на синтез продукции. Наибольшее количество теплоты образуется в органах с интенсивным обменом веществ и большой массой – печени и мышцах. При мышечной работе химическая энергия только на треть переходит в механическую работу, остальные две трети переходят в теплоту. Теплопродукция может увеличиваться в 3…5 раз за счет ...
0 комментариев