В. В. Шульговский
Московский государственный университет им. М.В. Ломоносова
Человек обладает совершенным зрением, и оно является важнейшим анализатором для познания окружающего мира. Зрение человека имеет высокое разрешение, оно цветное (приматы – трихроматы). Наш зрительный мир объемен. При движениях головы, ходьбе, беге, прыжках окружающий зрительный мир остается константным несмотря на то, что сетчаточное изображение испытывает сильные смещения. Из анатомии сетчатки известно, что колбочки – фоторецепторы дневного и цветного зрения – сосредоточены в желтом пятне (fovea), которое занимает на сетчатке всего около 2°. Именно они обеспечивают высокое разрешение зрения, восприятие цвета, предметное зрение. Зрение в сумерках, когда работают только палочки, занимающие остальное пространство сетчатки, чрезвычайно несовершенно – воспринимаются только контуры предметов (ночью все кошки серые). Какие нейрофизиологические механизмы обеспечивают совершенство нашего зрения кроме самих фоторецепторов? Чтобы разобраться в этом, обратимся к эволюции органов зрения в ряду позвоночных животных.
Степень развития зрения полностью определяется необходимостью животного ориентироваться во внешней среде. У рыб уже хорошо развито зрение. Сетчатка рыб в основных чертах не отличается от сетчатки млекопитающих. Аккомодация (наводка на резкость), правда, осуществляется с помощью передвижения круглого хрусталика ближе-дальше от сетчатки с помощью серповидноклеточного отростка, а не изменением кривизны хрусталика, как у наземных позвоночных. Уже в условиях однородной водной среды у рыб возникла необходимость специального механизма стабилизации изображения на сетчатке при движении животного. Голова рыбы непосредственно соединена с туловищем, и поэтому положение глаз в орбитах зависит от положения туловища. Движение глаз в орбитах определяется двумя рефлексами: вестибулоокулярным и оптомоторным.
Вестибулоокулярный рефлекс запускается с вестибулярных рецепторов, при этом глаза в орбитах противовращаются относительно направления вращения всего туловища. Благодаря этому рефлексу абсолютное положение зрительных осей в пространстве, окружающем животное, стабилизируется во время движения животного. Этот рефлекс для своей работы не требует зрительных стимулов. В отличие от этого оптомоторный рефлекс для своего запуска требует структурированного зрительного фона. В лаборатории оптомоторный рефлекс легко получить поместив животное в центре вращающегося цилиндра, стенки которого раскрашены черно-белыми полосами. Эти два рефлекса проявляются внешне одинаково: вестибулоокулярный рефлекс противовращает глаза в орбитах при вращении головы, сохраняя неизменным положение зрительных осей глаз в пространстве, а оптомоторный рефлекс также противовращает глаза в орбитах, но стабилизирует зрительные оси относительно структурированного зрительного поля зрения. Назначение обоих рефлексов – предотвратить сползание изображения при вращении животного или движении зрительного мира относительно животного.
Трудности в решении задачи стабилизации сетчаточного изображения многократно возрастают у наземных млекопитающих. Голова этих животных помещается на подвижной шее. Это дает дополнительные возможности исследовать окружающий мир, но в то же время резко усложняет проблему стабилизации сетчаточного изображения. Трудности стабилизации сетчаточного изображения возрастают, так как эти животные способны к быстрым движениям (бег, прыжки и пр.).
Эволюция сетчатки высших млекопитающих пошла по пути дифференцирования сетчатки на зону с высоким разрешением (area centralis – у копытных, хищных и некоторых других и fovea – у приматов), которая занимает, например, у человека около 2°. Остальная часть сетчатки, как уже упоминалось, занята палочками – рецепторами сумеречного зрения. В группе животных с area centralis (их условно можно назвать афовеальными) зрительная задача ограничивается прослеживанием движущихся объектов преимущественно по горизонтальному меридиану, поэтому движения глаз в орбитах ограничиваются плавными прослеживающими движениями. В группе фовеальных животных (в отряде приматов) возникла принципиально другая зрительная задача – активное исследование окружающего мира. Как уже говорилось, природа создала сетчатку приматов с fovea, в области которой находится высокая плотность колбочек – рецепторов дневного и цветового зрения. Для решения задачи исследования окружающего мира необходимо было создание глазодвигательного механизма активного помещения информативно значимых деталей сетчаточных изображений объектов на fovea (фовеация).
У человека наблюдаются все перечисленные выше рефлексы, при помощи которых осуществляются стабилизация изображения на сетчатке и рассматривание окружающего мира. Перечислим эти рефлексы. Вестибулоокулярный рефлекс легко получить вращением человека в специальном кресле, которое вращают вокруг вертикальной оси. Регистрация движений глазных яблок показывает, что при вращении человека в затемненной комнате глаза в орбитах испытывают колебательные движения (вестибулярный нистагм): медленная фаза движения в направлении вращения, а быстрая – в противоположную сторону. Например, этот рефлекс участвует в установке взора. Если предъявить зрительный объект на периферии зрительного поля, то человек совершает вначале скачкообразное движение (саккаду) глазами и тем самым помещает изображение на fovea. Но глаза в орбитах занимают в этом случае крайнее положение. Поэтому голова начинает поворачиваться в направлении зрительного объекта. Глаза при этом противовращаются в орбитах, удерживая изображение объекта на fovea. Благодаря всем этим движениям (глаз в орбитах и головы) положение зрительной оси в пространстве (мнимая линия, соединяющая fovea и зрительный объект) остается неизменным. Установка взора продолжается до тех пор, пока глаза в орбитах не занимают центрального положения. Оптомоторный рефлекс получают при движении перед глазами испытуемого чередующихся черно-белых полос. В этом случае движения глаз в орбитах не отличаются от вестибулоокулярного рефлекса – медленная фаза нистагма в направлении движения фона, а быстрая – в противоположную сторону. Рефлекс прослеживания хорошо проявляется при неожиданном появлении объекта в области бокового зрения. В этом случае глаз скачком приводит этот объект на центр сетчатки и затем осуществляются плавные прослеживающие движения глаз с участием головы. Совершенно уникальные движения глаз, которые в полной мере развиты только у приматов, включая человека, – это саккады (от фр. – хлопок паруса).
Современная нейрофизиология установила, что в производстве этой формы движения глаз задействован стволовой нейронный генератор. На сагиттальном срезе мозгового ствола головного мозга обезьяны показано местоположение области, ответственной за производство саккад, – она локализована в парамедианной области ретикулярной формации моста. Эта область содержит несколько видов нейронов. Управление этих нейронов производится из верхних двухолмий, а также эта область получает прямые входы от фронтального глазодвигательного поля коры больших полушарий. Благодаря последней связи управление саккадой может быть произвольным. Саккадными движениями глаз человек способен активно исследовать окружающий зрительный мир. Видно, что у больного человека нарушена произвольная способность управлять положением глаз и отчет таких больных не соответствует предъявленной картине.
Описанные глазодвигательные реакции чрезвычайно важны для зрительного восприятия. Например, расстройство вестибулоокулярного рефлекса, которое наблюдается при некоторых неврологических заболеваниях или в космическом полете у космонавтов, приводит к неспособности стабилизировать сетчаточное изображение и как следствие этого – к потере рассматриваемого объекта из поля зрения. Механизм саккад необходим для активного исследования окружающего мира, он принимает самое непосредственное участие в чтении, просмотре телевизионных передач и т.д. Перейдем к описанию некоторых свойств саккадной системы человека, связанных со зрительным вниманием.
Мы будем придерживаться трехуровневой гипотезы саккадной системы, которая предполагает три этапа в программировании саккады: процессы внимания, принятие решения и определение характеристик саккад. Эта идея получила название премоторной гипотезы внимания. Согласно этой гипотезе, механизм внимания можно разделить на процесс включения внимания, что может закончиться фовеацией, и сброса внимания. В результате последнего процесса информация поступает в систему локализации зрительной цели и происходит выбор объекта периферийным зрением. Сброс внимания может происходить при отсутствии зрительного восприятия. Опишем эксперимент, который привел к формулированию этой гипотезы.
Испытуемый в затемненной комнате находится перед экраном, в центре которого горит маленькая (около 2°) зрительная цель, так называемая фиксационная точка (ФТ). Испытуемый перед экспериментом получает инструкцию "фиксировать взор на светящейся точке". Если неожиданно для испытуемого погасить ФТ и зажечь другой стимул на расстоянии около 20° по горизонтальному меридиану – периферический стимул (ПС), то латентный период саккады на ПС будет составлять у здорового человека около 200-250 мс. Проведем этот эксперимент по другому – после гашения фиксационного стимула (ФС) следующий ПС появится только через 200 мс. Введение такого темнового зазора приводит к тому, что латентный период саккады на ПС сокращается в среднем на 40-60 мс. Был сделан вывод о том, что перевод взора с одной точки фиксации на другую состоит из следующих этапов: фиксация взора на ФС, освобождение от фиксации ФС – сброс внимания, вычисление параметров следующей саккады и перевод взора на ПС. Второй этап (сброс внимания с ФС) занимает 40-60 мс и может происходить в полной темноте.
Американский исследователь Мишель Познер рассматривает внимание как систему контроля умственных процессов, включающих три подсистемы: 1) ориентацию на сенсорное событие; 2) выделение зрительного сигнала для дальнейшей переработки и 3) поддержание активности головного мозга. Первые две подсистемы непосредственно связаны с движениями глаз. Внимание представляется как гипотетический луч и изучается его размер и характер распространения в зрительном поле. Интересно, что вертикальный меридиан, проходящий через fovea, оказался барьером на пути этого луча. Это проявляется в значительном увеличении латентного периода саккады или движения руки к стимулу, предъявляемому в неожиданной позиции в полуполе, противоположном фокусу внимания.
Экспериментальное исследование процессов зрительного внимания направлено на выявление мозговых структур, участвующих в этом процессе. Мы предполагаем, что этапам саккадического программирования могут соответствовать локальные изменения в биоэлектрических потенциалах различных корковых зон, которые могут быть выявлены с помощью метода картирования биопотенциалов. Этим методом выявляются структуры на всех уровнях головного мозга, которые составляют сложную иерархически параллельную систему произвольного управления саккадами у человека. В частности, большое число корковых полей больших полушарий непосредственно связано с этим процессом.
На рис. 5 показаны вызванные биоэлектрические потенциалы, которые регистрируются со скальпа здорового человека в условиях описанного выше эксперимента по предъявлению зрительных стимулов. Методом обратного усреднения биопотенциалов от начала саккады был выделен комплекс медленных и быстрых пресаккадических потенциалов. В период действия ФС на этапе ожидания ПС развивалась ранняя и поздняя медленная негативность (волны МН1 и МН2). В латентный период саккады были выделены быстрые негативные (N-100, N-200) и позитивные (P-100, P-200) потенциалы, а также спайковый (СП) потенциал, пик которого совпадал с началом саккады.
Картирование усредненных биопотенциалов в период центральной фиксации и межстимульной паузы в схеме направленного внимания показало, что ранняя премоторная негативность длительностью 300-500 мс локализована в передних лобных отделах, она симметрична и ее фокус активности переходит из Fz в F3 и F4. Он сильнее выражен перед коротколатентными саккадами, чем перед длиннолатентными, и в ситуации направленного внимания по сравнению с нейтральными условиями. По нашему мнению, это соответствует ориентировочному компоненту волны ожидания и отражает процессы внимания, мотивации, произвольной регуляции деятельности человека, связанные со зрительным восприятием. Центрально-лобная топография предполагает отражение в этом потенциале активности фронтомедиоталамической системы активации и срединных лимбических структур.
Поздняя медленная негативность преобладает в теменно-затылочных областях. В условиях вероятностного предъявления стимулов наблюдается переход фокусов из одного полушария в другое длительностью 50-70 мс. Фокус активности сильнее выражен перед длиннолатентными саккадами. Перед саккадами с коротким и средним латентным периодом фокус активности доминирует в контралатеральном к направлению движения глаз полушарии. Можно предположить, что такая динамика отражает процессы предварительной глазодвигательной подготовки, антиципации, указанных полей коры больших полушарий.
Таким образом, медленные негативные волны, развивающиеся при действии ПС в преднастроечный период, имеют различную пространственно-временную динамику и отражают участие в целенаправленном глазодвигательном поведении когнитивных процессов: внимания, ожидания и антиципации.
После включения целевого стимула в латентный период саккады развиваются быстрые пресаккадические потенциалы: P-200, N-200, P-100, N-100. Изучение параметров и топографии этих потенциалов в различных экспериментальных условиях позволило разделить их на две функциональные группы, отражающие этапы восприятия зрительных стимулов и программирования саккады. Ранние потенциалы P-200, N-200 коррелируют с процессами сенсорной переработки и "сброса внимания". Поздние потенциалы P-100, N-100 являются аналогами моторных потенциалов инициации саккад.
Как уже упоминалось, в схеме предъявления стимулов с темновым зазором в 200 мс наблюдают укорочение латентного периода саккады на 40-60 мс и появление большого числа так называемых экспресс-саккад с латентным периодом 90-120 мс. Феномен экспресс-саккад вызывает большой интерес и дискуссию. Наши экспериментальные данные свидетельствуют о включении в этот процесс дополнительного глазодвигательного поля, поля Fz. По-видимому, существует независимый путь в саккадический генератор мозгового ствола из Fz, минуя верхнее двухолмие. Это иллюстрирует рис. 6, на котором показаны усредненные вызванные потенциалы и их картирование перед экспресс-саккадой. Видно, что ПС включается на пике P-100. Картирование биопотенциалов показывает, что фокус активности находится в Fz, области дополнительного глазного поля. При высоком уровне внимания и функционального состояния головного мозга совпадение пространственного расположения стимула и предварительно выбранной из памяти цели вызывает, по нашему мнению, инициацию экспресс-саккады.
Таким образом, современные нейрофизиологические данные указывают на то, что в организации произвольных движений глаз человека задействовано большое число мозговых структур. На рис. 7 на схеме мозга человека отмечены основные структуры, участвующие в саккадической генерации (по характеристикам вызванных потенциалов). В ответ на предъявление зрительного сигнала вызванные потенциалы с латентным периодом в диапазоне 100-250 мс регистрируются в затылочных полях, а также в теменных и фронтальных полях. Подготовка к саккаде соответствует появлению поздней медленной негативности, которая регистрируется во фронтальном глазодвигательном поле и дополнительном глазодвигательном поле, а также в теменной коре. Процессы зрительного внимания коррелируют с ранней медленной негативностью. Судя по распространению этого потенциала, вовлекаются префронтальная кора, лимбическая кора, а также по данным на обезьянах интраламинарные ядра таламуса.
Современные нейрофизиологические исследования позволяют высказать гипотезу, что зрительное пространственное внимание является системой организации мозговых процессов, включающих ориентацию на зрительный стимул, выделение значимых элементов изображения для дальнейшей обработки, а также определяет зрительное восприятие.
Еще в прошлом веке В. Вундт указывал, что внимание можно определить как сфокусированное сознание на данном сигнале. Современная нейрофизиология позволяет определить некоторые пути экспериментального исследования сознания человека. Основная трудность, по-видимому, состоит в многозначности состояния сознания у человека (см. [1]). Частным случаем может быть восприятие сенсорного сигнала, которое включает как необходимый компонент процесс внимания. Экспериментальный подход, продемонстрированный в этой статье, позволяет современными нейрофизиологическими методами исследовать распределение фокусов возбуждения по коре больших полушарий головного мозга человека. В интерпретации нейрофизиологических данных представляется чрезвычайно плодотворной гипотеза, высказанная А.М. Иваницким. Он предполагает на основе экспериментальных данных, что процесс осознания связан с циркуляцией возбуждения и повторным его возвращением в корковые воспринимающие поля. Важным является также то, что этот возвратный сигнал должен быть дополнен также возбуждениями от лимбических структур головного мозга. Основные положения этой гипотезы хорошо прослеживаются при экспериментальном исследовании вызванных потенциалов головного мозга человека при произвольном восприятии зрительных сигналов. Именно процесс пространственного направленного зрительного внимания, которое мы создавали у испытуемого в нашем эксперименте, позволил исследовать большое число структур мозга, задействованных в этом психофизиологическом процессе зрительного восприятия.
Благодарю старшего научного сотрудника кандидата биологических наук М.В. Славуцкую за обсуждение статьи и ведущего научного сотрудника доктора биологических наук А.П. Кулаичева за техническое обеспечение экспериментальных исследований.
Список литературы
1. Симонов П.В. Лекции о работе головного мозга М.: ИП РАН, 1998.
2. Хрестоматия по вниманию / Под ред. А.Н. Леонтьева и др. М.: Изд-во МГУ, 1976.
3. Шульговский В.В. Физиология целенаправленного поведения млекопитающих. М.: Изд-во МГУ, 1993.
4. Posner M. Comulative development of attentional theory // Amer. Psychol. 1982. Vol. 37, Ъ 2. P. 168-176.
Похожие работы
... амплитуды ВП на то же стимул в покое, но без предварительного сигнала. Метод ВП создал новую эру в исследовании внимания, обеспечив исследователей новым инструментом для проверки теоретических схем. В свете накопленных в современной психологии и психофизиологии фактических данных и развиваемых представлений внимание следует рассматривать как один из процессов само регуляции, направленный на ...
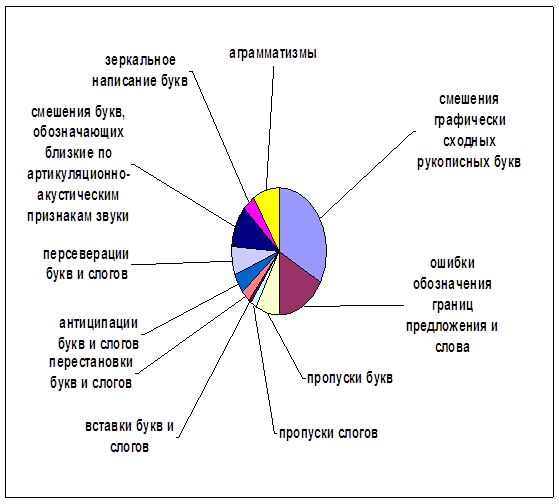

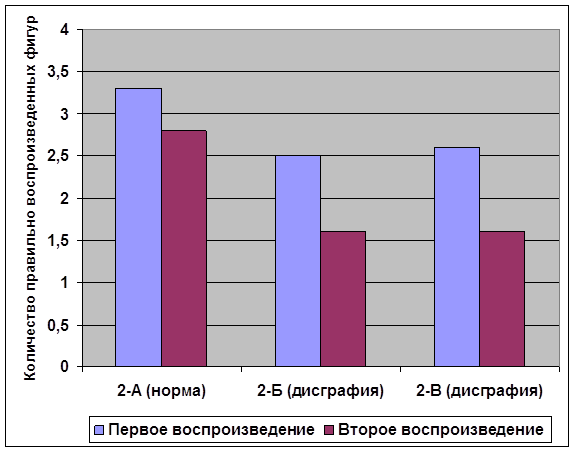
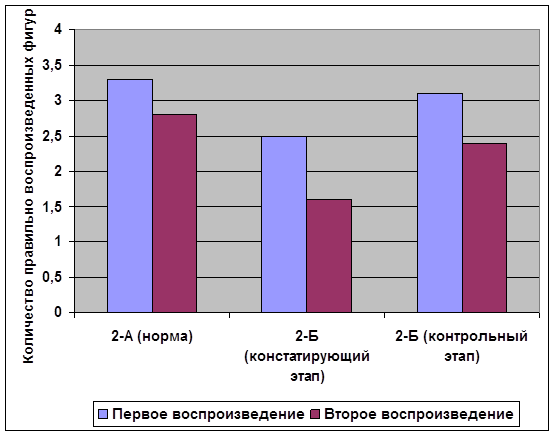
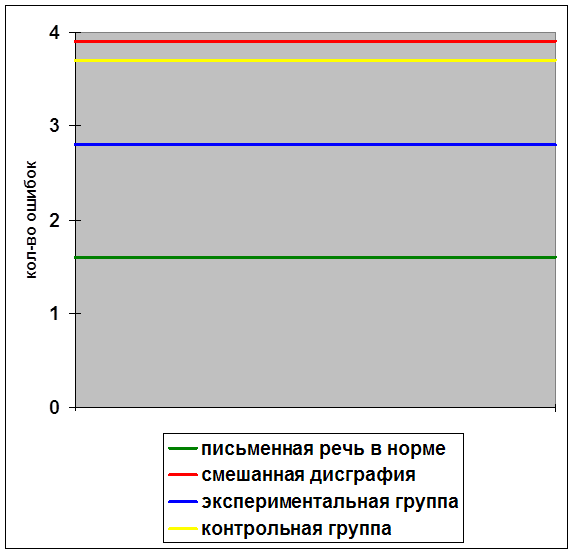
... внимания; развитие звукобуквенного анализа; развитие навыков работы с текстом. Для определения эффективности проведенной опытно-экспериментальной работы по развитию зрительной памяти в деле устранения смешанной дисграфии мы провели сравнительный анализ результатов, продемонстрированных детьми контрольной и экспериментальной групп, и сопоставительный анализ результатов детей экспериментальной ...

... ведется над преодолением нарушений оптического и оптико-пространственного гнозиса. 2. Экспериментальное изучение нарушений оптико – пространственных представлений у младших школьников и их коррекция 2.1. Выявление нарушений оптико – пространственных функций у младших школьников с оптической дисграфией Экспериментальное изучение проводилось с сентября по декабрь 2004 года на базе ...
... психологов нет единодушия во взглядах. Каждый из них рассматривает значение зрительных ощущений в различных аспектах: познавательных, адаптационных, развивающих, творческих. 1.2 Изучение проблемы зрительных ощущений в зарубежной психологии Иоганнес Мюллер выдвинул принцип специфических энергий чувств. основное положение, основной тезис Иоганнеса Мюллера. «Качество ощущения, — писал он, — ...
0 комментариев