Курсовая работа
Выполнила студентка 3-го курса ЭГО ЕГФ Гаганова В.В.
Липецкий государственный педагогический университет.
Естественно Географический Факультет.
Кафедра ботаники.
Липецк 2001.
Введение.
Вода – важнейший экологический фактор для всего живого на земле. Для процессов обмена веществ со средой, составляющих основу жизни, необходимо участие воды в качестве растворителя и метаболита. Так у растений вода участвует в реакциях фотосинтеза, минеральные соли поступают в растения из почвы только в виде водных растворов. Вода – главная составная часть тела растений. Даже находясь в анабиозе, растения содержат воду. Особая роль воды наземных растений заключается в постоянном пополнении больших трат ее на испарение в связи с развитием большой фотосинтезирующей поверхности. Вода, обуславливая необходимое тургорное давление, определенным образом участвует и в поддержании формы наземных растений как организмов не имеющих опорного скелета. Также для большой группы растений, живущих в водоемах, морях и океанах, вода является непосредственной средой обитания.
Глава I. Характеристика основных групп растений по отношению к воде.
По приуроченности к местообитаниям с разными условиями увлажнения и выработке соответствующих приспособлений среди наземных растений различают три основных экологических типа: гигрофиты, мезофиты и ксерофиты.
Растения, для которых вода не только необходимый экологический фактор, но непосредственная среда обитания, относятся к водным, называемым гидрофитами.
ГИГРОФИТЫ. Это растения избыточно увлажненных местообитаний с высокой влажностью воздуха и почвы. При довольно большом разнообразии местообитаний, особенностей водного режима и анатомо-морфологических черт всех гигрофитов объединяет отсутствие приспособлений, ограничивающих расход воды, и неспособность выносить даже незначительную ее потерю. Ярко выраженные гигрофиты – травянистые растения и эпифиты влажных тропических лесов, не выносящие сколько–нибудь заметного понижения влажности воздуха. Даже в разгар сезона дождей мелкие эпифитные папоротники на стволах деревьев теряют тургор и засыхают, если на них в течение 2-3 часов падают солнечные лучи. Черты гигрофитов имеют травянистые растения темнохвойных лесов (кислица, майник двулистный, двулепестник альпийский). К гигрофитам можно отнести и виды, растущие на открытых и хорошо освещенных местообитаниях, но в условиях избытка почвенной влаги – близ водоемов, в дельтах рек, в местах выхода грунтовых вод. В наших широтах примером могут служить прибрежные виды: калужница – Caltha palustris, плакун-трава – Lythrum salikaria, а в странах жаркого климата – папирус, болотные пальмы. Из культурных растений сюда можно отнести рис, культивируемый на полях, залитых водой.
КСЕРОФИТЫ. Это растения сухих местообитаний, способные переносить значительный недостаток влаги – почвенную и атмосферную засуху. Они распространены, обильны и разнообразны в областях с жарким и сухим климатом. К этой группе принадлежат виды пустынь, сухих степей, саванн, колючих редколесий, сухих субтропиков. В более гумидных районах ксерофиты участвуют в растительном покрове лишь в наиболее прогреваемых и наименее увлажненных местообитаниях (например, на склонах южной экспозиции).
Неблагоприятный водный режим растений в сухих местообитаниях обусловлен, во-первых, ограниченным поступлением воды при ее недостатке в почве и, во-вторых, увеличением расхода влаги на транспирацию при большой сухости воздуха и высоких температурах. Следовательно, для преодоления недостатка влаги возможны разные пути: увеличение ее поглощения и сокращение расхода, кроме того, способность переносить большие потери воды. Все это используется ксерофитами при адаптации к сухости, но у разных растений в неодинаковой степени, в связи с чем некоторые авторы различают два основных способа преодоления ксерофитами засухи: возможность противостоять иссушению тканей, или активное регулирование водного баланса, и способность выносить сильное иссушение.
В зависимости от структурных черт и способов регулирования водного режима различают несколько разновидностей ксерофитов (по Генкелю П.А.): эуксерофиты, гемиксерофиты, пойкилоксерофиты.
К группе ксерофитов относят и суккуленты – растения с сочными листьями или стеблями. Различают листовые суккуленты (агавы, алоэ) и стеблевые, у которых листья редуцированы, а наземные части представлены мясистыми стеблями(кактусы, некоторые молочаи).
Ксерофиты с наиболее ярко выраженными ксероморфными чертами строения листьев имеют своеобразный внешний облик, за что получили название склерофитов. Облик типичного склерофита легко представить на примере чертополоха – Carduus crispus и пустынных полыней, ковылей, саксаулов.
МЕЗОФИТЫ. Эта группа включает растения, произрастающие в средних условиях увлажнения. Сюда относятся растения лугов, травяного покрова лесов, лиственные древесные и кустарниковые породы из областей умеренно влажного климата, а также большинство культурных растений.
Мезофиты – группа весьма разнообразная не только по видовому составу, но и по различным экологическим оттенкам, обусловленным разным сочетанием факторов в природных местообитаниях. Они связаны переходами с другими экологическими типами растений по отношению к воде, так что четкую границу между ними провести очень трудно. Так, среди луговых мезофитов выделяются виды с повышенным влаголюбием, предпочитающие постоянно сырые или временно заливаемые участки (лисохвост луговой – Alopecurus pratensis, бекмания обыкновенная – Beckmannia eruciformis).
Их объединяют в переходную группу гигромезофитов наряду с некоторыми влаголюбивыми лесными травами, предпочитающими наиболее сырые леса, лесные овраги (недотрога – Impatiens nolitangere). С другой стороны в местообитаниях с переодическим или постоянным (небольшим) недостатком влаги много мезофитов с теми или иными ксероморфными признаками с повышенной физиологической устойчивостью к засухе. Эта группа переходная между мезофитами ксерофитами, - ксеромезофиты. Примером могут служить многие виды северных степей, сухих сосновых боров, песчаных местообитаний: клевер-белоголовка – Trifolium montanum, подмаренник желтый – Galium verum и другие.
Особое место среди мезофитов занимают степные и пустынные весенние эфемеры и эфемероиды. К этой группе принадлежат растения, ранней весной покрывающие степи и пустыни разноцветным цветущим ковром (многолетники – тюльпаны, гусиные луки; однолетники – маки, вероники). Это виды с чрезвычайно краткой вегетацией и длительным периодом покоя, который однолетние эфемеры переживают в виде семян, а многолетние эфемероиды – в виде покоящихся луковиц, клубней, корневищ. Кроме весенних существуют и осенние эфемероиды, произрастающие в районах с климатическим ритмом средиземноморского типа. Сюда относятся виды родов Crocus, Scilla и другие.
По многим особенностям структуры и физиологии близки к ксерофитам растения, которые по тем или иным причинам испытывают недостаток влаги, сопряженный с действием низких температур. Иногда такие виды в качестве особого подразделения включают в группу ксерофитов, иногда выделяют в самостоятельные экологические типы – психрофиты и криофиты.
Психрофиты – растения влажных и холодных почв в холодных местообитаниях высокогорий и северных широт. Несмотря на достаточное увлажнение почвы, они часто испытывают недостаток влаги (или из-за физиологической сухости, вызванной низкими температурами, или в связи с преобладанием в почве недоступной влаги, как, например, на торфянистых почвах). Среди психрофитов есть травянистые растения (например злаки северных лугов: белоус – Nardus strikta; высокогорные кавказские злаки: овсяница пестрая –Festuka varia), высокогорные, болотные и тундровые кустарники и кустарнички, как вечнозеленые (вереск – Calluna vulgaris), так и с опадающей листвой (карликовые ивы – Salix polaris, S. herbacea).К психрофитам относятся и хвойные древесные породы умеренных и северных широт.
Криофиты в экологическом отношении очень близки к психрофитам и связаны с ними переходными формами. Это растения сухих и холодных местообитаний – сухих участков тундр, скал, осыпей. Обычно они рассматриваются и характеризуются вместе с психрофитами, поскольку у них много сходных морфологических и физиологических черт. Но среди криофитов есть и весьма своеобразные формы – это растения-подушки высокогорных холодных пустынь.
ГИДРОФИТЫ. Это водные растения. По образу жизни и строению среди них можно выделить погруженные растения и растения с плавающими листьями. Погруженные растения подразделяют на укореняющиеся в донном грунте и взвешенные в толще воды. Из высших растений к первым принадлежат телорез – Stratiotes aloides, шильник водяной – Subularia aquatika. В эту же группу входят водоросли, прикрепленные к грунту. Из растений, взвешенных в толще воды, можно назвать роголистник погруженный—Ceratophyllum demersum, пузырчатку обыкновенную – Utrikularia vulgaris, а также многочисленные виды планктонных водорослей.
Растения с плавающими листьями используют частично водную, частично воздушную среду. Из них укореняются в грунте кувшинки из рода Nymphaea, кубышки из рода Nuphar, рдесты, орех водяной – Trapa natans.
Многие виды наряду с плавающими на поверхности воды листьями имеют и подводные. Плавают на поверхности воды, не укореняясь, ряски, водокрас.
К настоящим водным растениям очень близко примыкает и обычно вместе с ними рассматривается группа гелофитов или амфибий – земноводных растений. Это виды береговых и прибрежных местообитаний с избыточным или переменным увлажнением. Они могут расти как в воздушной среде, так и частично погруженными в воду, могут выносить и полное временное заливание. Как в природе нет резкой границы между водными и наземными местообитаниями для растений, так и группа гелофитов связана незаметными переходами, с одной стороны, с настоящими гидрофитами, с другой – с наземными гигрофитами и гигромезофитами. Примеры гелофитов – растений прибрежной полосы пресноводных водоемов и рек: стрелолист – Sagittaria sagittifolia, ежеголовка – Sparganium ramosum.
Глава II. Анатомо-морфологические приспособления растений к водному режиму.
Растения, приуроченные к местообитаниям с разными условиями увлажнения, выработали соответствующие приспособления к водному режиму.
Характерные структурные черты гигрофитов – тонкие нежные листовые пластинки с небольшим числом устьиц, не имеющие толстой кутикулы, рыхлое сложение тканей листа с крупными межклетниками, слабое развитие водопроводящей ткани, тонкие слаборазветвленные корни.
Для ксерофитов большое значение имеют разнообразные структурные приспособления к условиям недостатка влаги.
Корневые системы обычно сильно развиты, что помогает растениям увеличить поглощение почвенной влаги. По общей массе корневые системы ксерофитов нередко превышают надземные части, иногда весьма значительно. Так, у многих травянистых и кустарниковых видов среднеазиатских пустынь подземная масса больше надземной в 9-10 раз, а у кесрофитов памирских высокогорных холодных пустынь – в300-400 раз. Корневые системы ксерофитов часто бывают экстенсивного типа, то есть растения имеют длинные корни, распространяющиеся в большом объеме почвы, но сравнительно мало разветвленные. Проникновение таких корней на большую глубину позволяет ксерофитам использовать влагу глубоких почвенных горизонтов, а в отдельных случаях – и грунтовых вод.
У других видов корневые системы интенсивного типа: они охватывают относительно небольшой объем почвы, но благодаря очень густому ветвлению максимально используют почвенную влагу. Корни ряда ксерофильных видов имеют специальные приспособления для запасания влаги. Надземные органы ксерофитов также отличаются своеобразными (так называемыми ксероморфными чертами), которые носят отпечаток трудных условий водоснабжения. У них сильно развита водопроводящая система, что хорошо заметно по густоте сети жилок в листьях, подводящих воду к тканям. Эта черта облегчает ксерофитам пополнение запасов влаги, расходуемой на транспирацию.
Разнообразные структурные приспособления защитного характера, направленные на уменьшение расхода воды, в основном сводятся к следующему:
Общее сокращение транспирирующей поверхности. Многие ксерофиты имеют мелкие, узкие, сильно редуцированные листовые пластинки. В особо засушливых пустынных местообитаниях листья некоторых древесных и кустарниковых пород редуцированы до едва заметных чешуек. У таких видов фотосинтез осуществляют зеленые ветви.
Уменьшение листовой поверхности в наиболее жаркие и сухие периоды вегетационного сезона. Для многих кустарников среднеазиатских, североафриканских и других пустынь, а так же для некоторых видов сухих субтропиков средиземноморья характерен сезонный деформизм листьев: ранней весной при еще благоприятном водном режиме образуются относительно крупные листья, которые летом, при наступлении жары и сухости, сменяются мелкими листьями более ксероморфного строения с меньшей интенсивностью транспирации.
Защита листьев от больших потерь влаги на транспирацию. Она достигается благодаря развитию мощных покровных тканей – толстостенного, иногда многослойного эпидермиса, часто несущего различные выросты и волоски, которые образуют густое “войлочное” опушение поверхности листа. У других видов поверхность покрыта водонепроницаемым слоем толстой кутикулы или воскового налета. Развитие защитных покровов на листьях причина того, что степной травостой имеет тусклые, седоватые оттенки, резко отличающиеся от яркой зелени лугов.
Устьица у ксерофитов обычно защищены от чрезмерной потери влаги, например, расположены в специальных углублениях в ткани листа, иногда снабженных волосками и прочими дополнительными защитными устройствами. У ковылей и других степных злаков существует интересный механизм защиты устьиц в самые жаркие и сухие часы дня: при больших потерях воды крупные тонкостенные водоносные клетки эпидермиса теряют тургор, и лист свертывается в трубку; так устьица оказываются изолированными от окружающего сухого воздуха внутри замкнутой полости, где благодаря транспирации создается повышенная влажность. Во влажную погоду клетки эпидермиса восстанавливают тургор, и листовая пластинка вновь развертывается.
Усиленное развитие механической ткани.
Клетки тканей листьев у ксерофитов отличаются мелкими размерами и весьма плотной упаковкой, то есть малым развитием межклетников, благодаря чему сильно сокращается внутренняя испаряющая поверхность листа. Поскольку ксерофиты обычно обитают на открытых, хорошо освещенных местообитаниях, многие черты ксероморфной структуры листа – это одновременно и черты световой структуры. Так у многих видов листья имеют мощную иногда многорядную палисадную паренхиму, часто расположенную с обеих сторон.
Основные морфолого-анотомические черты мезофитов – средние между чертами гигрофитов и ксерофитов. Мезофиты имеют умеренно развитые корневые системы как экстенсивного, так и интенсивного типа, со всеми переходами между ними. Для листа характерна дифференцировка тканей на более или менее плотную палисадную паренхиму и рыхлую губчатую паренхиму с системой межклетников. Сеть жилок сравнительно негустая. Покровные ткани могут иметь отдельные ксероморфные черты, но не столь ярко выраженные, как у ксерофитов.
Психрофиты имеют ярко выраженную ксероморфную структуру листа. Так, психрофильные злаки узколистны, имеют хорошо развитую проводящую и механическую ткани; некоторые из них способны к свертыванию листовой пластинки в трубку, напоминая степные ксерофиты. Вечнозеленые кустарнички имеют плотные кожистые листья, иногда с весьма мощной кутикулой, плотной палисадной паренхимой. Нижняя сторона, несущая устьица, часто опушена или покрыта восковым налетом. Вместе с тем в структуре листа ясно заметны и некоторые гигроморфные черты, главным образом, крупные размеры клеток и развитие больших межклетников в губчатой ткани.
Анатомо-морфологические черты гидрофитов существенно отличают их от наземных растений. Заметна редукция механических тканей. Столь же сильно редуцированна и проводящая система. Если у сухопутных мезофитов длина жилок на 1 кв.см. листа составляет около100 мм и более, а у ксерофитов доходит до300, то у водных и прибрежных растений она в несколько раз меньше. Вот несколько примеров (по Гесснеру Ф., 1959):
| Вид | Плавающие листья, мм | Подводные листья, мм |
| Nelumbium nuciferum | 71 | — |
| Victoria cruciana | 41 | — |
| Victoria regia | 20 | — |
| Potamogeton praelongus | — | 14 |
У некоторых погруженных растений, не прикрепленных к грунту, корни полностью редуцированны, другие корни сохранили, но отдельно плавающие части растений могут обходиться и без них. Корни укрепляющихся гидрофитов слабоветвистые, без корневых волосков. Вместе с тем ряд видов имеет толстые и прочные корневища, которые играют роль якоря, хранилища запасных веществ и органа вегетативного размножения.
Листья погруженных гидрофитов очень тонки и нежны, имеют упрощенное строение мезофилла без заметной дифференциации на палисадную и губчатую паренхиму. Подводные листья без устьиц. В отдельных местах находятся группы клеток эпидермиса с утонченными стенками. Считается, что они играют большую роль в поглощении воды и растворенных минеральных солей.
У растений лишь частично погруженных в воду, хорошо выражена гетерофиллия - различие строения надводных и подводных листьев на одной и той же особи. Первые имеют черты, обычные для листьев наземных растений, вторые – очень тонкие или рассеченные листовые пластинки. Гетерофиллия отмечена у водного лютика – Ranunculus diversifolius, кувшинок и кубышек, стрелолиста и других видов. Интересный пример – поручейник, на стебле которого можно видеть несколько форм листьев, представляющих все переходы от типично наземных до водных.
Глава III.Физиологические адаптации растений, приуроченных к местообитаниям разной увлажненности.
Наряду с морфологическими особенностями у растений, приуроченных к местам с разными условиями увлажненности, выработались и физиологические.
Способность гигрофитов к регуляции водного режима ограничена: устьица большей частью широко открыты, так что транспирация мало отличается от физического испарения. Благодаря беспрепятственному потоку воды и отсутствию защитных приспособлений интенсивность транспирации очень высока: у световых гигрофитов в дневное время листья могут терять за час количество воды, в 4-5 раз превышающее массу листа. Высокая оводненность тканей гигрофитов поддерживается в основном за счет постоянного притока влаги из окружающей среды.
Другие характерные физиологические черты гигрофитов, обусловленные легкой доступностью влаги, - низкое осмотическое давление клеточного сока, незначительная водоудерживающая способность, приводящая к быстрой потере запасов воды.особенно показательны для гигрофитов небольшие величины сублетального водного дефицита: так, для кислицы и майника потеря 15%-20% запаса воды уже необратима и ведет к гибели. В некоторых случаях у растений сильно увлажненных местообитаний возникает необходимость удаления избытка влаги. Обычно, это бывает, когда почва хорошо прогрета и корни активно всасывают воду, а транспирация отсутствует (например, утром или при тумане, когда влажность воздуха 100%). Избыточная влага удаляется путем гуттации – выделение воды через специальные выделительные клетки, расположенные по краю или на острие листа.
Ксерофиты обладают рядом разнообразных физиологических адаптаций, позволяющих им успешно выдерживать недостаток влаги.
У ксерофитов обычно повышено осмотическое давление клеточного сока, позволяющее всасывать воду даже при больших водоотнимающих силах почвы, то есть использовать не только легкодоступную, но и труднодоступную почвенную влагу. Оно измеряется тысячами кПа, а у некоторых пустынных кустарников зарегистрированы цифры, достигающие 10000-30000 кПа.
С давних пор пристальное внимание привлекала проблема расхода воды ксерофитами на транспирацию. Казалось бы, многочисленные анатомические приспособления, достаточно надежно защищающие наземные части ксерофитов от сильного испарения, должны способствовать значительному снижению транспирации. Однако выяснилось, что в действительности это не так. При достаточном водоснабжении большинство ксерофитов имеют довольно высокую транспирацию, но при наступлении засушливых условий, они сильно сокращают ее. При этом играет роль и закрывание устьиц, и сильное обезвоживание листа при начинающемся подвядании. Несомненно, анатомо-морфологические приспособления имеют определенное значение, но основную роль в засухоустойчивости ксерофитов в настоящее время отводят физиологическим механизмам.
К числу этих механизмов принадлежит высокая водоудерживающая способность тканей и клеток, обусловленная рядом физиологических и биохимических особенностей.
Большое значение для выживания ксерофитов при резком недостатке влаги имеет их способность переносить глубокое обезвоживание тканей без потери жизнеспособности и способности восстановления нормального содержания воды в растении при возобновлении благоприятных условий. Ксерофиты способны потерять до 75% всего водного запаса и, тем не менее, остаться живыми. Ярким примером в этом отношении служат пустынные растения, которые летом высыхают до состояния, близкого к воздушно-сухому, и впадают в анабиоз, но после дождей возобновляют рост и развитие.
Еще одна система адаптаций, обеспечивающих выживание ксерофитов в аридных условиях, - выработка сезонных ритмов, дающих возможность растениям использовать для вегетации наиболее благоприятные периоды года и резко сократить жизнедеятельность во время засухи. Так, в областях со средиземноморским климатом с резко выраженным летним сухим периодом многие ксерофильные виды имеют “двухтактный” ритм сезонного развития: весенняя вегетация сменяется летним покоем, во время которого растения сбрасывают листву и снижают интенсивность физиологических процессов; в период осенних дождей вегетация возобновляется, и затем уже следует зимний покой. Сходное явление наблюдается и у растений сухих степей в середине и конце лета: потеря части листовой поверхности, приостановка развития, сильное обезвоживание тканей и т.д. Такое состояние, получившее название полупокоя, длится вплоть до осенних дождей, после которых у степных ксерофитов начинают отрастать листья.
Физиологические адаптации суккулентов столь своеобразны, что их необходимо рассмотреть отдельно.
Основной способ преодоления засушливых условий у суккулентов – накопление больших запасов воды в тканях и крайне экономное ее расходование. В условиях жаркого и сухого климата весь водный запас мог бы быть быстро растрачен, но растения имеют защитные приспособления, направленные к сокращению транспирации. Одно из них – своеобразная форма надземных частей суккулентов. В дополнение к этому у многих суккулентов поверхность защищена восковым налетом опушением, хотя есть и суккуленты с тонким не защищенным эпидермисом. Устьица очень немногочисленны, часто погружены в ткань листа или стебля. Днем устьица обычно закрыты, и потеря воды идет в основном через покровные ткани.
Транспирация у суккулентов чрезвычайно мала. Ее трудно уловить за короткий период и приходится определять расход воды не за час, а за сутки или за неделю. Водоудерживающая способность тканей суккулентов значительно выше, чем у других растений экологических групп, благодаря содержанию в клетках гидрофильных веществ. Поэтому и без доступа влаги суккуленты расходуют водный запас очень медленно и долго сохраняют жизнеспособность даже в гербарии.
Ограничения, обусловленные особенностями водного режима суккулентов, создают и другие трудности для жизни этих растений в аридных условиях. Слабая транспирация сводит к минимуму возможность терморегуляции, с чем связано сильное нагревание массивных надземных органов суккулентов. Затруднения создаются и для фотосинтеза, поскольку днем устьица обычно закрыты, а открываются ночью, следовательно, доступ углекислоты и света не совпадают во времени. Поэтому у суккулентов выработался особый путь фотосинтеза, при котором в качестве источника углекислоты, частично используются продукты дыхания. Иными словами, в крайних условиях растения частично используют принцип замкнутой системы с реутилизацией отходов метаболизма.
В силу всех этих ограничений интенсивность фотосинтеза суккулентов невелика, рост и накопление массы идут очень медленно, вследствие чего они не отличаются высокой биологической продуктивностью и не образуют сомкнутых растительных сообществ.
Физиологические показатели водного режима мезофитов подтверждают их промежуточную позицию: для них характерны умеренные величины осмотического давления, содержания воды в листьях, предельного водного дефицита. Что касается транспирации, то ее величина в большей степени зависит от условий освещенности и других элементов микро климата.
Один и тот же мезофильный вид, попадая в разные по водоснабжению условия, обнаруживает известную пластичность, приобретая в сухих условиях более ксероморфные, а во влажных более гигроморфные черты.
Пластичность листьев проявляется не только в разных местообитаниях, но даже у одной и той же особи. Например, у деревьев на опушке леса листья на стороне, обращенной в сторону леса, имеют более мезофильный и теневой характер по сравнению с несколько ксероморфными листьями внешней стороны дерева (см. табл. ниже ). Листья разных высотных ярусов одних и тех же растений находятся в неодинаковых условиях водоснабжения, так как поступление воды в верхние части связано с преодолением большого сопротивления. К тому же у деревьев верхние листья обычно находятся в условиях иного микроклимата.
Различие анатомо-физиологических показателей листьев на разных сторонах кроны дерева, растущего на опушке леса.
I – сторона, обращенная к лесу, II – сторона, обращенная к поляне
| Древесная порода | Площадь листа, см2 | Число устьиц на 1 мм2 | Содержание воды, % | Содержание хлорофилла, мг/г | Средняя интенсивность фотосинтеза, мг СО2 /дм2 ·ч | Толщина листа, мкм |
| Дуб – Quercus robur | ||||||
| I | 42 | 45 | 61 | 3.0 | 1.9 | 97 |
| II | 18 | 125 | 54 | 2.4 | 2.3 | 181 |
| Липа – Tilia cordata | ||||||
| I | 38 | 38 | 71 | 3.6 | 1.6 | 93 |
| II | 24 | 45 | 62 | 2.0 | 1.1 | 106 |
Водная среда существенно отличается от воздушной, поэтому у водных растений существует ряд своеобразных физиологических адаптивных черт. Интенсивность света в воде сильно ослаблена, поскольку часть падающей радиации отражается от поверхности воды, другая – поглощается ее толщей. В связи с ослаблением света фотосинтез у погруженных растений сильно снижается с увеличением глубины. Считают, сто выживанию глубоководного фитопланктона в зонах, где освещенность ниже точки компенсации, способствуют его периодические вертикальные перемещения в верхние зоны, где идет интенсивный фотосинтез и пополнение запасов органических веществ.
В воде кроме недостатка света растения могут испытывать и другое затруднение, существенное для фотосинтеза, - недостаток доступной СО2 . Углекислота поступает в воду в результате растворения СО2 , содержащегося в воздухе, дыхания водных организмов, разложения органических остатков и высвобождения из карбонатов. При интенсивном фотосинтезе растений идет усиленное потребление СО2 , в связи с чем легко возникает ее дефицит.
На увеличение содержания СО2 в воде гидрофиты реагируют заметным повышением фотосинтеза.
У погруженных растений транспирации нет, значит, нет и “верхнего двигателя”, поддерживающего ток воды в растении. Однако этот ток, доставляющий к тканям питательные вещества, существует, при чем с явной суточной периодичностью: днем больше, ночью отсутствует. Активная роль в его поддержании принадлежит корневому давлению и деятельности специальных клеток, выделяющих воду, - водяных устьиц.
Плавающие или торчащие над водой листья обычно имеют сильную транспирацию, хотя и расположены в слое воздуха, который непосредственно граничит с водой и имеет повышенную влажность. Устьица широко открыты и закрываются полностью только в ночное время.
Столь же велика транспирация у прибрежных растений, при чем у них значительное количество воды расходуется не только листьями, но и стеблями.
Осмотическое давление у водных и прибрежных растений очень низкое, так как им не приходится преодолевать водоудерживающую силу почвы при поглощении воды.
Примеры осмотического давления (в кПа) у водных растений и гелофитов (по Гесснеру Ф.,1959)
| Название растения | Давление в корне | Давление в листе |
| Водные растения | ||
| Hippuris vulgaris | 940 | 1120 |
| Nymphaea dentata | 580 | 760 |
| Victoria regia | 580 | 760 |
| Гелофиты | ||
| Polygonum amphibium | — | 950 |
| Menyanthes trifoliata | 580 | 1120 |
| Lythrum salicaria | — | 1190 |
| Typha angustifolia | — | 880 |
Список литературы
Ботаника с основами экологии: Учеб. Пособие для студентов пед. ин-тов /Л.В. Кудряшов и др. – М.: Просвещение,1979
Горышина Т.К. Экология растений. – М.: Высшая школа,1979
Гусев Н.А. Некоторые закономерности водного режима растений. – М.: Изд-во АН СССР, 1959.
Двораковский М.С. Экология растений. – М.: Просвещение 1964
Жданов В.С. Аквариумные растения. / Под. ред. Коровина. - М.: Лесн. пром-ть , 1981
Крафтс А. Вода и ее значение в жизни растений. – М.: Иност. лит., 1951
Культиасов И.М. Экология растений. - М.: Изд-во московского ун-та, 1982
Мак-Дуголл В.Б. Экология растений. – М.: Учпедгиз, 1935.
Пономарева И.Н. Экология растений с основами биогеоценологии. – М.: Просвещение, 1978
Рычин Ю.В. Флора гигрофитов. Определитель. – М.: Сов. наука, 1948
Шенников А.П. Экология растений. – М.: Сов. наука, 1950
Похожие работы
... , часть накопленных в процессе фотосинтеза углеводов теряется. Однако все попытки искусственно ингибировать фотодыхание приводили к общему снижению интенсивности фотосинтеза. На современном этапе развития физиологии растений принято считать, что основное значение фотодыхания заключается в его защитной роли. Сбрасывая таким образом избыточную энергию, растение избегает разрушения фотосистем, ...
... . У засухоустойчивых растений эти показатели будут выше. В селекционной работе используют такой показатель, как содержание статолитного крахмала в корневом чехлике. Генетически обусловленным признаком засухоустойчивости растений является способность их вегетативных органов (особенно листьев) накапливать во время засухи пролин. При этом концентрация пролина увеличивается в 10—100 раз. В пролине ...
... и гибридов тех или иных сельскохозяйственных культур может различаться и по качеству урожая – содержанию белка, клейковины, крахмала, сахара, жира и т.д. Однако при агроэкологической оценке сельскохозяйственных культур в конкретных почвенно-климатических условиях необходимо использовать региональные справочные материалы, что позволит свести к минимуму ошибки в оптимизации структуры посевных ...
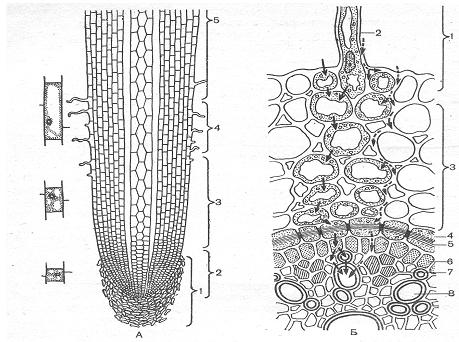
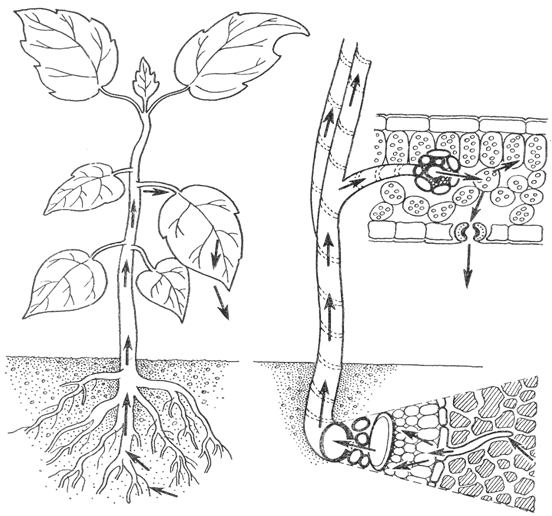
... , опыты показали, что вода может поступать в побеги и через мертвую корневую систему, причем в этом случае поглощение воды даже ускоряется. Кроме верхнего концевого двигателя водного тока, в растениях существует нижний концевой двигатель. Это хорошо доказывается на примере таких явлениях, как гуттация. Листья растений, клетки которых насыщены водой, в условиях высокой влажности воздуха, ...
0 комментариев