Раздражимость - это общее свойство тканей реагировать на различные раздражители.
Возбудимость - более узкое понятие, которое характеризует свойство тканей возбуждаться в ответ на действие раздражителя. Ткани, обладающие этим свойством, называются возбудимыми. Проявляется возбуждение возникновением потенциала действия. В основе возбуждения лежат сложные физико-химические процессы. Начальный пусковой момент возбуждения - изменения ионной проницаемости и электрических потенциалов мембраны. Возбудимые ткани имеют ряд свойств: раздражимость - способность тканей воспринимать раздражение, возбудимость - способность тканей реагировать возбуждением на раздражение, проводимость - способность распространять возбуждение, лабильность - скорость протекания элементарных циклов возбуждения. Лабильность отражает время, в течение которого ткань восстанавливает работоспособность после очередного цикла возбуждения. Порог раздражения (в физиологии нервных и мышечных клеток), наименьшая сила раздражителя (обычно электрического тока), способная вызвать распространяющийся потенциал действия
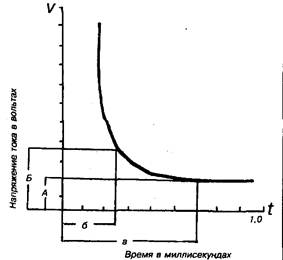
Методы изучения описанных явлений разнообразны. Так, о возбудимости можно судить по наименьшей силе раздражителя, необходимой для возникновения той или иной рефлекторной реакции или по пороговой силе тока или пороговому сдвигу потенциала, достаточным для возникновения ПД. Здесь необходимо ввести такие понятия, как реобаза и хронаксия. Реобаза (от греч. rheos - течение, поток и basis - ход, движение; основание), наименьшая сила постоянного электрического тока, вызывающая при достаточной длительности его действия возбуждение в живых тканях. Понятие реобазы и хронаксии ввёл в физиологию Л. Лапик в 1909, определяя зависимость между силой тока и длительностью его действия при изучении наименьшего (порогового) эффекта возбудимых тканей. Реобаза, как и хронаксия, даёт представление о возбудимости тканей и органов по порогу силы и длительности действия раздражения. Реобаза соответствует порогу раздражения и выражается в вольтах или миллиамперах. Значение реобазы можно вычислить по формуле: i = a/t + b, где i - сила тока, t - длительность его действия, а и b - константы, определяемые свойствами ткани. Константа b является Р., так как при длительном действии раздражающего тока отношение a/t будет очень мало и i практически равняется b. Р. нередко называются пороговые значения не только электрических, но и других раздражителей. Хронаксия (от греч. chronos - время и axia - цена, мера), наименьшее время действия на ткань постоянного электрического тока удвоенной пороговой силы (удвоенной реобазы), вызывающего возбуждение ткани. Было также экспериментально установлено (голландский физик Л. Горвег, 1892, французский физиолог Ж. Вейс, 1901), что величина стимула, вызывающего возбуждающий эффект в тканях, находится в обратной зависимости от длительности его действия и графически выражается гиперболой - кривая <сила - время. Минимальная сила тока, которая при неограниченно долгом действии вызывает эффект возбуждения (реобаза), соответствует на рисунке отрезку OA (BC). Наименьшее т. н. полезное время действия порогового раздража
ющего стимула соответствует отрезку OC (полезное потому, что дальнейшее увеличение времени действия тока не имеет значения для возникновения потенциала действия). При кратковременных раздражениях кривая силы - времени становится параллельной оси ординат, т. е. возбуждение не возникает при любой силе раздражителя. Приближение кривой асимптотически к линии, параллельной абсциссе, не позволяет достаточно точно определять полезное время, т.к. незначительные отклонения реобазы, отражающие изменения функционального состояния биологических мембран в покое, сопровождаются значительными колебаниями времени раздражения. В связи с этим Лапик предложил измерять другую условную величину - хронаксию, т. е. время действия раздражителя, равное двойной реобазе [на рисунке соответствует отрезку OD (EF)]. При данной величине раздражителя наименьшее время его действия, при котором возможен пороговый эффект, равно OF. Установлено, что форма кривой, характеризующей возбудимость ткани в зависимости от интенсивности и длительности действия раздражителя, однотипна для самых разнообразных тканей. Различия между ними касаются только абсолютного значения соответствующих величин и, прежде всего, времени, т. е. возбудимые ткани отличаются друг от друга временной константой раздражения. Лабильность можно измерить, раздражая ткань электрическим током различной частоты. Момент, когда ткань произойдёт преобразование ритма (ткань перестанет воспроизводить заданный ритм без изменений) и будет лабильностью данной ткани. Единицы её измерения - количество воспроизводимых импульсов за единицу времени [имп./сек.(мин.), и т. д. ]. Проводимость можно охарактеризовать расстоянием, преодолённым импульсом за единицу времени, то есть скоростью распространения импульса.
История вопроса.Изучался этот вопрос давно. Многих учёных интересовал тот факт, что некоторые животные способны генерировать <электрический ток. Это явление было названо <животное электричество. К этому времени относится начало исследований итальянского физиолога и врача Л. Гальвани, заложивших основу учения о Б. п. Многолетний научный спор (1791-97) между Л. Гальвани и физиком А. Вольта о природе <животного электричества завершился двумя крупными открытиями: были получены факты о существовании биоэлектрических явлений в живых тканях и открыт новый принцип получения электрического тока с помощью разнородных металлов - создан гальванический элемент (вольтов столб). Правильная оценка наблюдений Гальвани стала возможной лишь после применения достаточно чувствительных электроизмерительных приборов - гальванометров. Первые такие исследования были проведены итальянским физиком К. Маттеуччи (1837). Систематическое изучение биопотенциалов было начато немецким физиологом Э. Дюбуа-Реймоном (1848), который доказал существование биопотенциалов в нервах и мышцах в покое и при возбуждении. Но ему не удалось (в силу большой инерционности гальванометра) зарегистрировать быстрые, длящиеся тысячные доли секунды колебания биопотенциалов при проведении импульсов вдоль нервов и мышц. В 1886 немецкий физиолог Ю. Бернштейн проанализировал форму потенциала действия; французский учёный Э. Ж. Марей (1875) применил для записи колебаний потенциалов бьющегося сердца капиллярный электрометр; русский физиолог Н. Е. Введенский использовал (1883) для прослушивания ритмических разрядов импульсов в нерве и мышце телефон, а голландский физиолог В. Эйнтховен (1903) ввёл в эксперимент и клиническую практику струнный гальванометр - высокочувствительный и малоинерционный прибор для регистрации электрических токов в тканях. Значительный вклад в изучение биопотенциалов внесли русские физиологи: В. В. Правдич-Неминский (1913-21) впервые зарегистрировал электроэнцефалограмму, А. Ф. Самойлов (1929) исследовал природу нервно-мышечной передачи возбуждения, а Д. С. Воронцов (1932) открыл следовые колебания биопотенциалов, сопровождающие потенциал действия в нервных волокнах. Дальнейший прогресс в изучении биопотенциалов был тесно связан с успехами электроники, позволившими применить в физиологическом эксперименте электронные усилители и осциллографы (работы американских физиологов Г. Бишопа, Дж. Эрлангера и Г. Гассера в 30-40-х гг. 20в.). Изучение биопотенциалов в отдельных клетках и волокнах стало возможным с разработкой микроэлектродной техники. Важное значение для выяснения механизмов генерации биопотенциалов имело использование гигантских нервных волокон головоногих моллюсков, главным образом кальмара. Диаметр этих волокон в 50 - 100 раз больше, чем у позвоночных животных, он достигает 0,5-1 мм, что позволяет вводить внутрь волокна микроэлектроды, инъецировать в протоплазму различные вещества и т.п. Изучение ионной проницаемости мембраны гигантских нервных волокон позволило английским физиологам А. Ходжкину, А. Хаксли и Б. Катцу (1947-52) сформулировать современную мембранную теорию возбуждения.
Потенциал покоя. Потенциал действия.Потенциал покоя (ПП) - разность потенциалов между содержимым клетки (волокна) и внеклеточной жидкостью; скачок потенциала локализуется на поверхностной мембране, при этом её, внутренняя сторона заряжена электроотрицательно по отношению к наружной. Потенциал покоя обусловлен неравенством концентраций, ионов Na+, К+ и Cl- по обе стороны клеточной мембраны и неодинаковой её проницаемостью для этих ионов. В нервных и мышечных клетках потенциал покоя участвует в поддержании состояния готовности молекулярной структуры мембраны к возбуждению в ответ на действие раздражителя. Все воздействия на клетку, вызывающие длительное стойкое снижение потенциала покоя. (например, нарушение обмена веществ, повышение внеклеточного содержания ионов К+, действие сильного электрического тока и т.д.), ведут к снижению возбудимости клетки или к полной утрате ею способности к генерации потенциалов действия. У живых клеток в покое между внутренним содержимым клетки и наружным раствором существует разность потенциалов (ПП) порядка 60-90мв, которая локализована на поверхностной мембране. Внутренняя сторона мембраны заряжена электроотрицательно по отношению к наружной.
Концентрация К+ в протоплазме примерно в 50 раз выше, чем во внеклеточной жидкости, поэтому, диффундируя из клетки, ионы выносят на наружную сторону мембраны положительные заряды, при этом внутренняя сторона мембраны, практически не проницаемой для крупных органических анионов, приобретает отрицательный потенциал. Поскольку проницаемость мембраны в покое для Na+ примерно в 100 раз ниже, чем для К+, диффузия натрия из внеклеточной жидкости (где он является основным катионом) в протоплазму мала и лишь незначительно снижает ПП, обусловленный ионами К+. В скелетных мышечных волокнах в возникновении потенциала покоя важную роль играют также ионы Cl-, диффундирующие внутрь клетки. Следствием ПП является ток покоя, регистрируемый между поврежденным и интактным участками нерва или мышцы при приложении отводящих электродов. Мембраны нервных и мышечных клеток (волокон) способны изменять ионную проницаемость в ответ на сдвиги мембранного потенциала. При увеличении ПП (гиперполяризация мембраны) проницаемость поверхностных клеточных мембран для Na+ и К+ падает, а при уменьшении ПП (деполяризация) она возрастает, причём скорость изменений проницаемости для Na+ значительно превышает скорость увеличения проницаемости мембраны для К+. Неравенство концентраций ионов К+ и Na+ (или Ca+) внутри и снаружи клетки (волокна) поддерживается специальным механизмом (т. н. <натриевым насосом), выталкивающим ионы Na+ из клетки и нагнетающим ионы К+ в протоплазму, требующим затраты энергии, которая черпается клеткой в процессах обмена веществ. Работа таких механизмов обеспечивается, как правило, энергией, выделяемой при расщеплении аденозинтрифосфорной кислоты (АТФ); таким образом, ионные насосы одновременно выполняют функцию ферментов, расщепляющих АТФ и называемых АТФ-азами. Активный перенос Na+ из клетки сопряжён с транспортом К+ в обратном направлении и осуществляется особой ферментной системой - транспортной Na, К, - стимулируемой аденозинтрифосфатазой, локализованной в клеточной мембране. Последняя, гидролизуя аденозинтрифосфорную кислоту (АТФ), высвобождает энергию, которая и затрачивается на активный перенос катионов. Работа насоса в целом зависит от уровня метаболизма клетки.
Потенциал действия.Потенциал действия (ПД). Все раздражители, действующие на клетку, вызывают в первую очередь снижение ПП; когда оно достигает критического значения (порога), возникает активный распространяющийся ответ - ПД. Во время восходящей фазы ПД кратковременно извращается потенциал на мембране: её внутренняя сторона, заряженная в покое электроотрицательно, приобретает в это время положительный потенциал. Достигнув вершины, ПД начинает падать (нисходящая фаза ПД), и потенциал на мембране возвращается к уровню, близкому к исходному, - ПП. Полное восстановление ПП происходит только после окончания следовых колебаний потенциала - следовой деполяризации или гиперполяризации, длительность которых обычно значительно превосходит продолжительность пика ПД. Согласно мембранной теории, деполяризация мембраны, вызванная действием раздражителя, приводит к усилению потока Na+ внутрь клетки, что уменьшает отрицательный потенциал внутренней стороны мембраны - усиливает её деполяризацию. Это, в свою очередь, вызывает дальнейшее повышение проницаемости для Na+ и новое усиление деполяризации и т.д. В результате такого взрывного кругового процесса, т. н. регенеративной деполяризации, происходит извращение мембранного потенциала, характерное для ПД. Повышение проницаемости для Na+ очень кратковременно и сменяется её падением (рис. 3), а следовательно, уменьшением потока Na+ внутрь клетки. Проницаемость для К+, в отличие от проницаемости для Na+, продолжает увеличиваться, что приводит к усилению потока К+ из клетки. В результате этих изменений ПД начинает падать, что ведёт к восстановлению ПП. Таков механизм генерации ПД в большинстве возбудимых тканей. Существуют, однако, клетки (мышечные волокна ракообразных, нервные клетки у ряда брюхоногих моллюсков, некоторые растительные клетки), у которых восходящая фаза ПД обусловлена повышением проницаемости мембраны не для ионов Na+, а для ионов Ca+. Своеобразен также механизм генерации ПД в мышечных волокнах сердца, для которых характерно длительное плато на нисходящей фазе ПД.
Ионные механизмы потенциала действия.В нервных волокнах восходящая фаза ПД связана с активацией т. н. быстрых натриевых каналов (БНК), а нисходящая фаза - с инактивацией БНК и активацией калиевых каналов (КК). На таком же механизме основана генерация ПД в волокнах скелетных мышц позвоночных. В мышечных волокнах сердца активация БНК обеспечивает только начальный подъём ПД Характерное же для этих волокон плато ПД связано с активированием медленных натрий-кальциевых каналов (МНК).
Изучение физико-химических свойств ионных каналов важно не только для расшифровки их молекулярной структуры, но и для разработки методов управления генерацией ПД в различных клетках. Установлено, что БНК специфически блокируются тетродотоксином (ядом японской рыбы-шар и калифорнийских саламандр), а также новокаином, кокаином и др. местными анестезирующими средствами. МНК и МКК к этим агентам нечувствительны, но блокируются ионами Mn2+, Со2+, Ni2+, La3+ и органическими соединениями - изоптином (используемым в кардиологической практике) и его дериватом Д-600. Большинство КК эффективно блокируется тетраэтиламмонием. Пусковое влияние ПД на такие внутриклеточные процессы, как сокращение миофибрилл (в скелетных, гладких и сердечной мышцах), нейросекреция (в некоторых специализированных нейронах и нервных окончаниях) и т.д., осуществляется в результате прямого воздействия электрического импульса; на внутриклеточные структуры (выброс) ионов Ca2+ из саркоплазматической сети мышцы) и влияния на эти структуры ионов Ca2+, проникающих внутрь клетки во время ПД.
Изменение возбудимости ткани при её возбуждении.Фазовые колебания потенциала действия влияют на возбудимость клетки. Изменения возбудимости также имеют фазовый характер и находятся в тесной взаимосвязи с ПД.
Максимальная возбудимость клетки соответствует фазе ПП. Рассмотрим Схему соотношения ПД и возбудимости. При деполяризации мембраны возбудимость повышается (фаза латентного дополнения) (1). После достижения мембранным потенциалом критического уровня деполяризации происходит овершут. В этот момент возбудимость практически мгновенно падает до нуля. Это - фаза абсолютной рефрактерности (2). Ни один импульс, пришедший в эту фазу не способен возбудить клетку. По мере восстановления мембранного потенциала (реполяризации) восстанавливается и возбудимость (фаза относительной рефрактерности) (3). В эту фазу надпороговые раздражители способны возбудить клетку. Эта фаза обусловлена повышением проницаемости мембраны для ионов K+, который выходит из цитоплазмы, снижая заряд мембраны внутри клетки. В фазу следовой деполяризации возбудимость несколько превышает нормальную - фаза экзальтации (4). Однако, поскольку K+ - каналы медленные, то калий выходит из клетки даже в несколько избыточном количестве, что ведёт к возникновению гиперполяризации. Возбудимость при этом несколько снижается (фаза субнормальной возбудимости) (5). После этого мембранный потенциал приходит к исходному значению, восстанавливается и возбудимость (6).
Похожие работы
... ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ (два занятия) Занятие 1-е ПРИРОДА ВОЗБУЖДЕНИЯ 1.Что называют раздражимостью и возбудимостью? 2.Каково соотношение понятий раздражимость и возбудимость? Какие ткани в физиологии называют возбудимыми,какие- невозбудимыми? 3.Клетки каких тканей организма являются возбудимыми и невозбудимыми? 4.Дайте определение понятию "раздражитель". 5.Назовите два вида ...
ают под миогенным механизмом регуляции.Перечислите органы,для которых этот вид регуляции является важным. 6.Перечислите основные особенности гуморальной регуляции функций. 7.Перечислите особенности нервной регуляции по сравнению с гуморальной. 8.Назовите виды влияний нервной системы на органы,поясните их сущность. 9.При ...
... это противоположные по значению саморегулирующиеся процессы, которые устанавливают "золотую середину" уровня отношений организма со средой. Нервные волокна (отростки нервных клеток) обладают всеми свойствами возбудимых тканей, а проведение нервных импульсов является их специальной функцией. Скорость проведения возбуждения зависит от: 1 - диаметра волокон (толще ® быстрее), 2 - строения их ...
... концевой пластинки). Вызванный потенциал – это потенциал действия нейрона, возникающий в ответ на возбуждение рецептора, несущего информацию к этому нейрону. История исследования физиологии возбуждения Л. Гальвани был первым, кто убедился в существовании «живого электричества». Его первый (балконный) опыт состоял в том, что препарат задних лапок лягушек на медном крючке был подвешен к ...
0 комментариев