Реферат на тему:
ОСНОВНЫЕ ГЕНЕТИЧЕСКИЕ
ХАРАКТЕРИСТИКИ ПОПУЛЯЦИИ
Москва, 2001 г.
Популяция (франц. population – население) – совокупность особей одного вида, занимающих определенный ареал, свободно скрещивающихся друг с другом, имеющих общее происхождение, генетическую основу и в той или иной степени изолированных от других популяций данного вида.
Популяция – элементарная эволюционная структура. Основу современной эволюционной теории, которую называют неодарвинизмом или синтетической теорией эволюции, составляет изучение популяционной генетики. Гены, действуя независимо или совместно с факторами среды, определяют фенотипические признаки организмов и обуславливают изменчивость в популяциях. Фенотипы, приспособленные к условиям данной среды или «экологическим рамкам», сохраняются отбором, тогда как неадаптивные фенотипы подавляются и, в конце концов, элиминируются. Естественный отбор, влияя на выживание отдельных особей с данным фенотипом, тем самым определяет судьбу их генотипа, однако лишь общая генетическая реакция всей популяции определяет выживание данного вида, а также образование новых видов. Только те организмы, которые, прежде чем погибнуть успешно произвели потомство, вносит вклад в будущее своего вида. Для истории данного вида судьба отдельного организма не имеет существенного значения.
ГЕНОФОНДГенетически популяция характеризуется ее генофондом (аллелофондом).
Генофонд представлен совокупностью аллелей, образующих генотипы организмов данной популяции. В каждой данной популяции состав генофонда из поколения в поколение может постоянно изменяться. Новые сочетания генов образуют уникальные генотипы, которые в своем физическом выражении, т. е. в форме фенотипов, подвергаются давлению факторов среды, производящих непрерывный отбор и определяющих, какие гены будут переданы следующему поколению.
Популяция, генофонд которой непрерывно изменяется из поколения в поколение, претерпевает эволюционное изменение. Статичный генофонд отражает отсутствие генетической изменчивости среди особей данного вида и отсутствие эволюционного изменения.
Генофонды природных популяций отличает наследственное разнообразие (генетическая гетерогенность, или полиморфизм), генетическое единство, динамическое равновесие доли особей с разными генотипами.
Наследственное разнообразие заключается в присутствии в генофонде одновременно различных аллелей отдельных генов. Первично оно создается мутационным процессом. Мутации, будучи обычно рецессивными и не влияя на фенотипы гетерозиготных организмов, сохраняются в генофондах популяций в скрытом от естественного отбора состоянии. Накапливаясь, они образуют резерв наследственной изменчивости. Благодаря комбинативной изменчивости этот резерв используется для создания в каждом поколении новых комбинаций аллелей. Объем такого резерва огромен. Так, при скрещивании организмов, различающихся по 1000 локусов, каждый из которых представлен десятью аллелями, количество вариантов генотипов достигает 101000.
Генетическое единство популяции обуславливается достаточным уровнем панмиксии. В условиях случайного подбора скрещивающихся особей источником аллелей для генотипов организмов последовательных поколений является весь генофонд популяций. Генетическое единство проявляется также в общей генетической реализации популяции при изменении условий существования, что обуславливает как выживание вида, так и образование новых видов.
ЗАКОН ХАРДИ – ВАЙНБЕРГАЛюбой физический признак определяется одним или несколькими генами. Каждый ген может существовать в нескольких различных формах – аллелях. Число организмов в данной популяции, несущих определенный аллель, определяет частоту данного аллеля (которую иногда называют частотой гена, что менее точно). Например, у человека частота доминантного аллеля, определяющего нормальную пигментацию кожи, волос и глаз, равна 99%. Рецессивный аллель, детерминирующий отсутствие пигментации – так называемый альбинизм, - встречается с частотой 1%. В популяционной генетике частоту аллелей или генов выражают не в процентах, а в десятичных дробях. Таким образом, в данном случае частота доминантного аллеля равна 0,99 , а частота рецессивного аллеля альбинизма – 0,01. Общая частота аллелей в популяции составляет 100%, или 1,0.
Как это принято в классической генетике, аллели обозначаются буквами, например A – доминантный аллель, a – рецессивный и рецессивный) в генофонде популяции, используются символы p и q. Таким образом,
![]()
где p – частота доминантного, q – частота рецессивного аллеля. (В примере с пигментацией у человека p = 0,99 , а q =0,01). Значение этого уравнения состоит в том, что, зная частоту одного из аллелей, можно определить частоту другого.
Частоты отдельных аллелей в генофонде позволяют вычислять генетические изменения в данной популяции и определять частоту генотипов.
Математическая зависимость между частотами аллелей и генотипов в популяциях была установлена в 1908 г. независимо друг от друга английским математиком Дж. Харди и немецким врачом В. Вайнбергом. Эта зависимость получила название закон Харди – Вайнберга (равновесие Харди – Вайнберга). Закон этот гласит: «В бесконечно большой популяции из свободно скрещивающихся особей в отсутствие мутаций, избирательной миграции организмов с различными генотипами и давления естественного отбора первоначальные частоты доминантного и рецессивного аллелей сохраняются постоянными из поколения в поколение».
Поэтому любые изменения частоты аллелей должны быть обусловлены нарушением одного или нескольких перечисленных выше условий. Все эти нарушения способны вызвать эволюционное изменение. Эти изменения и их скорость можно изучить и измерить с помощью уравнения Харди – Вайнберга.
Если имеется два организма, один гомозиготный по доминантному аллелю A, а другой – по рецессивному аллелю a, то все их потомки будут гетерозиготными

Если наличие доминантного аллеля A обозначить символом p, а рецессивного аллеля a – символом q, то картину скрещивания между особями F1, возникающие при этом генотипы и их частоты можно представить следующим образом:
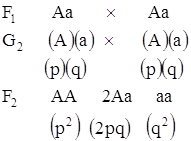
Используя символы p и q результаты приведенного выше скрещивания можно представить следующим образом:
p2 – доминантные гомозиготы
2pq – гетерозиготы
q2 – рецессивные гомозиготы.
Такое распределение возможных генотипов носит статистический характер и основано на вероятностях. Три возможных генотипа, образующихся при таком скрещивании, представлены со следующими частотами:
AA 2Aa aa0,25 0,50 0,25
Сумма частот трех генотипов, представленных в рассматриваемой популяции, равна единице; пользуясь символами p и q, можно сказать, что вероятности генотипов следующие:
![]()
На математическом языке ![]() представляет собой уравнение вероятности, тогда как
представляет собой уравнение вероятности, тогда как ![]() является квадратом этого уравнения (т.е.
является квадратом этого уравнения (т.е. ![]() ).
).
Поскольку
p – частота доминантного аллеля;
q – частота рецессивного аллеля;
p2 – гомозиготный доминантный тип;
2pq – гетерозиготный генотип;
q2 – гомозиготный рецессивный генотип,
можно вычислить частоты всех аллелей и генотипов, пользуясь выражениями
для частот аллелей: ![]()
для частот генотипов: ![]()
Однако, для большинства популяций частоту обоих аллелей можно вычислить только по доле особей, гомозиготных по рецессивному аллелю, так как это единственный генотип, который можно распознать по его фенотипическому выражению.
Например, один человек из 10000 – альбинос, т.е. частота альбинотического генотипа составляет 1 на 10000. Поскольку аллель альбинизма рецессивен, альбинос должен быть гомозиготным по рецессивному гену, т.е. на языке теории вероятности
![]()
Зная, что q2 =0,0001, можно определить частоты аллеля альбинизма (q), доминантного аллеля нормальной пигментации (p), гомозиготного доминантного генотипа (p2) и гетерозиготного генотипа (2pq). Так как q2=0,0001,
![]() ,
,
т.е. частота аллеля альбинизма в популяции равна 0,01 или 1%. Поскольку
![]()
![]()
частота доминантного аллеля в популяции равна 0,99 или 99%. А если
![]() и
и ![]() , то
, то
![]() ,
,
т.е. частота гетерозиготного генотипа составляет 0,0198; иными словами, примерно 2% индивидуумов в данной популяции несут аллель альбинизма либо в гетерозиготном либо в гомозиготном состоянии.
Гетерозиготных индивидуумов, нормальных по фенотипу, но обладающих рецессивным геном, который в гомозиготном состоянии может вызвать нарушение метаболизма, называют носителями. Существование в популяции неблагоприятных аллелей в составе гетерозиготных генотипов называют генетическим грузом. Как показывают вычисления с использованием уравнения Харди – Вайнберга, частота носителей в популяции всегда выше, чем можно было бы ожидать на основании фенотипического проявления данного дефекта.
СЛЕДСТВИЯ УРАВНЕНИЯ ХАРДИ – ВАЙНБЕРГАИз уравнения Харди – Вайнберга следует, что значительная доля имеющихся в популяции рецессивных аллелей находится у гетерозиготных носителей. Фактически гетерозиготные генотипы служат важным потенциальным источником генетической изменчивости. Это приводит к тому, что в каждом поколении из популяции может элиминироваться лишь очень малая доля рецессивных аллелей. Только те рецессивные аллели, которые находятся в гомозиготном состоянии, проявятся в фенотипе и тем самым подвергнутся селективному воздействию факторов среды и могут быть элиминированы. Многие рецессивные аллели элиминируются потому, что они неблагоприятны для фенотипа – обуславливают либо гибель организма еще до того как он успеет оставить потомство, либо «генетическую смерть», т.е. неспособность к размножению.
Однако не все рецессивные аллели неблагоприятны для популяции. Например, у человека из всех групп крови чаще всего встречается группа 0, соответствующая гомозиготности по рецессивному аллелю.
Другим примером служит серновидноклеточная анемия. наследственное заболевание, широко распространенное в ряде областей Африки, индии, у негритянского населения Северной Америки. Индивидуумы, гомозиготные по соответствующему рецессивному аллелю, обычно умирают не достигнув половой зрелости и элиминируя таким образом из популяции по два рецессивных аллеля. Что касается гетерозигот, то они не гибнут. Установлено, что во многих частях земного шара частота аллеля серновидноклеточности остается относительно стабильной. У некоторых африканских племен частота гетерозиготного фенотипа достигает 40%. Раньше думали, что этот уровень поддерживается за счет появления новых мутантов. Однако в результате дальнейших исследований выяснилось, что во многих частях Африки, где среди факторов, угрожающих здоровью и жизни, важное место занимает малярия, люди, несущие аллель серновидноклеточности, обладают повышенной резистентностью к этой болезни. В малярийных районах Центральной Африки это селективное преимущество гетерозиготного генотипа поддерживает частоту аллеля серновидноклеточности на уровне 10 – 20%. У североамериканских негров, уже 200 – 300 лет не испытывающих на себе селективного эффекта малярии, частота аллеля серновидноклеточности упала до 5%.
Этот пример эволюции в действии ясно демонстрирует селективное влияние среды на частоту аллелей – механизм нарушающий равновесие Харди – Вайнберга. Именно такого рода механизмы вызывают в популяциях сдвиги, ведущие к эволюционному изменению.
ФАКТОРЫ, ВЫЗЫВАЮЩИЕ ИЗМЕНЕНИЯ
В ПОПУЛЯЦИЯХ
Принцип Харди – Вайнберга носит чисто теоретический характер. Очень немногие популяции находятся в условиях, при которых сохраняется равновесие.
Половая рекомбинация (кроссинговер во время мейоза, независимое распределение хромосом при мейозе, случайное оплодотворение) обуславливает перетасовку генов, лежащую в основе происходящих непрерывных изменений. Но хотя эти процессы и приводят к образованию новых генотипов и изменяют их частоты, они не вызывают никакого изменения имеющихся аллелей, так что частоты аллелей остаются постоянными.
А эволюционные изменения происходят вслед за появлением новых аллелей, а главным источником последних служат мутации.
МУТАЦИОННЫЙ ПРОЦЕССБольшой вклад в популяционную генетику внес русский ученый С. С. Четвериков, обративший внимание на насыщенность природных популяций рецессивными мутациями, а также на колебания частоты генов в популяциях в зависимости от действия факторов внешней среды.
Мутационный процесс, изменяя частоту одного аллеля по отношению к другому, оказывает на генофонд популяции прямое действие. За счет мутантных аллелей происходит формирование резерва наследственной изменчивости. Благодаря мутационному процессу поддерживается высокий уровень наследственного разнообразия природных популяций. Совокупность аллелей, возникающих в результате мутаций, составляет элементарный эволюционный материал.
Большинство мутаций первоначально оказывает на фенотип особей неблагоприятное действие. В силу рецессивности мутантные аллели обычно присутствуют в генофондах популяций в гетерозиготных по соответствующему локусу генотипах.
Благодаря этому достигается тройственный положительный результат: 1) исключается непосредственное отрицательное влияние мутантного аллеля на фенотипическое выражение признака, контролируемого данным геном; 2) сохраняются нейтральные мутации, не имеющие приспособительной ценности в настоящих условиях существования, но которые смогут приобрести такую ценность в будущем; 3) накапливаются некоторые неблагоприятные мутации, которые в гетерозиготном состоянии нередко повышают относительную жизнеспособность организмов (эффект гетерозиса). Таким образом, создается резерв наследственной изменчивости.
НЕСЛУЧАЙНОЕ СКРЕЩИВАНИЕВ большинстве природных популяций спаривание происходит неслучайным образом. Во всех тех случаях, когда наличие одного или нескольких наследуемых признаков повышает вероятность успешного оплодотворения гамет, имеет место половой отбор. У растений и животных существует много структурных и поведенческих механизмов, исключающих чисто случайный подбор родительских особей. Например, цветки, у которых лепестки крупнее и нектара больше, чем обычно, вероятно будут привлекать больше насекомых, что повысит вероятность опыления и оплодотворения. Характер окраски насекомых, рыб и птиц и особенности их поведения, связанные с постройкой гнезда, охраной территории и брачными церемониями, повышают избирательность при скрещивании.
Таким образом, половой отбор как механизм избирательного скрещивания обеспечивает некоторым особям более высокий репродуктивный потенциал, в результате чего вероятность передачи генов этих особей следующему поколению повышается. Репродуктивный потенциал особей с менее благоприятными признаками понижен, и передача их аллелей последующим поколениям происходит реже.
ДРЕЙФ ГЕНОВО дрейфе генов говорят в тех случаях, когда изменение частоты генов в популяциях бывают случайными и не зависят от естественного отбора. Случайный дрейф генов или генетико-автоматические процессы, или эффект Сьюэлла Райта, может служить важным механизмом эволюционных изменений в небольших и изолированных популяциях. В небольшой популяции могут быть представлены не все аллели, типичные для данного вида.
Генетический дрейф может возникнуть при отделении новой популяции от родительской из-за того, что процент определенных аллелей среди отделившихся особей будет иным, чем в исходной популяции. Например, некий аллель встречается в популяции в 1% случаев. Допустим, что 10 животных из этой популяции были занесены на изолированный остров и там образовали новую популяцию. Если среди этих десяти одно было носителем данного аллеля, то его частота в новой популяции будет составлять уже 10%. Если же не было ни одного, то в образовавшейся популяции этот аллель будет отсутствовать.
Или же, например, если данный аллель встречался в популяции из 1000000 особей с частотой, допустим 1% (т.е. q = 0,01), то им будет обладать 10000 особей; а в популяции состоящей из 100 особей, этот аллель будет иметься только у одной особи, так что вероятность его случайной утраты в малой популяции (например, в результате преждевременной гибели особи – единственного носителя данного аллеля в популяции) будет гораздо выше.
С дрейфом генов связано явление, известное под названием принципа основателя. Оно состоит в том, что при отделении от родительской популяции небольшой ее части последняя может случайно оказаться не вполне типичной по своему аллельному составу. Некоторые аллели в ней могут отсутствовать, а другие будут представлены с непропорционально высокой частотой. Постоянное скрещивание внутри такой пионерной популяции приведет к созданию генофонда, отличающегося по частотам аллелей от генофонда исходной родительской популяции. Дрейф генов обычно снижает генетическую изменчивость в популяции, главным образом в результате утраты тех аллелей, которые встречаются редко. Суть дрейфа генов в том, что из-за случайных колебаний один из аллелей может вообще выпасть из популяции и она окажется по данному гену гомозиготной. В результате происходит гомозиготизация особей и затухание изменчивости. Это открывает путь случайной дифференцировке популяций вплоть до видового обособления.
ПОТОК ГЕНОВПоток генов – это перемещение аллелей из одной популяции в другую в результате скрещивания между членами этих двух популяций. Случайное внесение новых аллелей в популяцию – реципиента и удаление их из популяции – донора изменяет частоту аллелей в обеих популяциях и ведет к повышению генетической изменчивости. Несмотря на то, что поток генов вносит в популяции генетическую изменчивость, в смысле эволюционного изменения его действие оказывается консервативным. Распространяя мутантные аллели по всем популяциям, поток генов приводит к тому, что все популяции данного вида приобретают общий генофонд, т.е. различия между популяциями уменьшаются.
_______________________________
Таким образом, изменение частоты генов, вызванные теми или иными факторами внешней среды, служат основой возникновения различий между популяциями и в дальнейшем обуславливают преобразование их в новые виды. Поэтому изменения популяций в ходе естественного отбора называют микроэволюцией.
ЛИТЕРАТУРА
1. Грин Н., Стаут У., Тейлор Д. «Биология» в 3-х тт. М. 1993
2. Чебышев Н.В., Кузнецов С.В., Зайчикова С.Г., Барабанов Е.И. «Эволюция и экология». М. 1995
3. Мамонтов С.Г., Захаров В.Г. «Общая биология». М. 1996
4. Слюсарев А.А. «Биология с общей генетикой». М. 1979
5. Каменский А.А., Соколова Н.А., Титов С.А. «Биология. Ответы на вопросы». М. 1998
6. «Биология» под ред. проф. Ярыгина В.Н. М. 1997
7. Пуговкин А.П., Пуговкина Н.А. «Основы общей биологии». СПб. 1995
Похожие работы

... невозможно. Поэтому вводятся соответствующие обобщенные показатели для всех людей, позволяющие затем в среднем определить соотношение генетического и средового влияния на отдельного индивида. Изучение генеалогическим методом семей лиц, страдающих психическими болезнями, убедительно показало накопление в них случаев психозов и аномалий личности. Увеличение частоты случаев болезни среди близких ...
... т.д. Зная эти закономерности, можно количественно исследовать генетическую структуру популяций и прогнозировать ее возможные изменения. Этому способствует солидная теоретическая база популяционной генетики – популяционно-генетические процессы математически формализованы и описаны уравнениями динамики. Для проверки различных гипотез о генетических процессах в популяциях разработаны статистические ...
... также общностью заселяемой территории. Типичным примером популяции в животноводстве является порода. Сохранение и усиление хозяйственно-полезных признаков в популяции происходит благодаря получению нового поколения. Чем короче интервал между поколениями, тем быстрее будут получены животные с желательными признаками и тем быстрее можно будет достичь нужного результата. Достижение поставленной цели ...
... особи. Зародилось и успешно функционирует целое научное направление – демэкология (от греч. demos – народ), рассматриваемое как раздел общей экологии, изучающий структурные и функциональные характеристики, динамику численности популяции, внутрипопуляционные группировки и их взаимоотношения, выясняющий условия, при которых формируются популяции, и др. При этом приоритетной для демэкологии ...
0 комментариев