МИНИСТЕРСТВО ОБЩЕГО И ПРОФЕССИОНАЛЬНОГО ОБРАЗОВАНИЯ
РОССИЙСКОЙ ФЕДЕРАЦИИ
НИЖЕГОРОДСКИЙ ОРДЕНА ТРУДОВОГО КРАСНОГО ЗНАМЕНИ
ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ ИМ. Н.И.ЛОБАЧЕВСКОГО
БИОЛОГИЧЕСКИЙ ФАКУЛЬТЕТ
КАФЕДРА БИОФИЗИКИ
Реферат на тему
Методы измерения ионных токов
Выполнил студент
4 курса гр.144-7
Савин А.В.
Нижний Новгород
1998
1. Метод фиксации потенциала
Для
изучения
потенциалзависимых
мембранных
каналов применяется
метод фиксации
потенциала.
В данном методе
используют
электронную
систему с обратной
связью, которая
обеспечивает
автоматическое
поддержание
мембранного
потенциала.
Разность потенциалов
по разные стороны
мембраны фиксируют
на определенном
уровне, при
этом мембранный
потенциал можно
ступенчато
изменят на
строго определенную
величину. Такой
метод позволяет
измерить ионные
токи, протекающие
сквозь мембрану
через каналы,
которые активируются
при изменении
потенциала.
В соответствии
с законом Ома,
если напряжение
на мембране
постоянно,
изменения тока
однозначно
связанные с
изменениями
проводимости.
В свою очередь,
мы можем фиксировать
мембранный
потенциал на
разном уровне
и измерять
возникающие
при этом токи.
Если же использовать
растворы с
различенным
ионным составом,
и препараты,
избирательно
блокирующие
тот или иной
канал, то можно
будет изучать
поведение
различных
ионных каналов,
через которые
протекают
измеряемые
нами токи. Технически
фиксация потенциала
осуществляется
следующим
образом. При
помощи усилителя-регулятора
внутриклеточный
потенциал
сравнивают
с управляющим
потенциалом
(см. рис.1). Любое
отклонение
мембранного
потенциала
от управляющего
усиливается
и на выходе
усилителя
возникает
управляющий
ток. Этот ток
течет через
электроды,
расположенные
по разные стороны
мембраны в
таком направлении,
что мембранный
потенциал вновь
становится
равным управляющему.
Такое автоматическое
согласование
происходит
за долю миллисекунды
после того, как
задается ступенчатый
у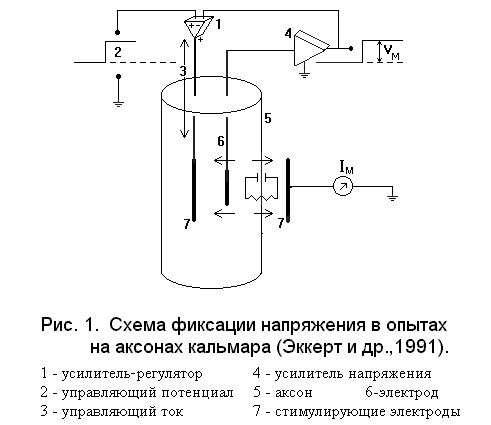
правляющий
потенциал.
Когда в ответ на такую ступенчатую деполяризацию открываются натриевые (или какие-либо другие) каналы, соответствующие ионы входят в аксон по электрохимическому градиенту и переносят с собой электрические заряды. Эти входящие заряды стремятся сдвинуть мембранный потенциал в положительном направлении, однако малейшее отклонение от управляющего потенциала немедленно компенсируется в результате удаления из клеток избыточных зарядов с помощью усилителя-регулятора. При этом записывается тот ток, который подается усилителем для поддержания мембранного потенциала на необходимом уровне, и этот ток в точности равен ионному току, протекающему через мембрану.
Метод фиксации напряжения, или кламп метод, позволяет измерять ионный поток при перемещении иона по контролируемому градиенту электрохимического потенциала и получать информацию об электрической проводимости мембраны и её пассивной проницаемости в отношении интересующего нас иона. Этот метод используется для измерения вольт-амперных характеристик растительных клеток, что позволяет получить информацию о тонких механизмах функционирования различных систем мембранного транспорта.
2. Метод пэтч-кламп
Метод пэтч-кламп (patch-clamp) позволяет осуществлять локальную (точечную) фиксацию мембранного потенциала и измерять токи через одиночные ионные каналы. На данный момент этот метод является мощным средством для исследования биомембран. Метод позволяет:
проводить многие исследования в рамках классических электрофизиологических подходов.
регистрировать токи и потенциалы от клеток очень малых размеров (3-10 мкм)
регистрировать токи одиночных каналов амплитудой порядка пикоампер
исследовать действие лекарственных препаратов при быстром подведении их как к наружной, так и к внутренней стороне мембраны
Метод пэтч-кламп был введен в исследовательскую практику Неером и Сакманом, когда в 1976 году ими была опубликована статья в журнале “Nature”, которая называлась “Токи через одиночные каналы в мембране волокна денервированной мышцы лягушки”. Это открыло путь для изучения на молекулярном уровне электрических свойств мембран и регуляции различных транспортных процессов.
Основой для создания метода послужило обнаружение факта, что при определённых условиях клеточная мембрана формирует очень плотный контакт с поверхностью кончика стеклянного микроэлектрода. При небольшом разрежении, создаваемом внутри пипетки, между стеклом и мембранным фрагментом возникает контакт, имеющий гигаомное сопротивление. В результате образуется электрически изолированный участок мембраны, и шум регистрирующего сигнала уменьшается на несколько порядков. Так как контакт мембраны со стеклом очень прочен, то находящийся под кончиком электрода фрагмент надо либо изолировать от клетки, либо разрушить, и таким образом проникнуть внутрь клетки. Существует несколько вариантов метода пэтч-кламп (рис.2).
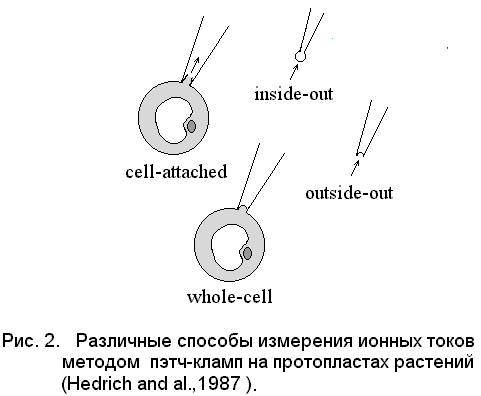
Наиболее близким к естественным условиям является вариант измерения ионных токов на прикреплённой, но неповрежденной (cell-attached) клетке, поскольку исследуемый участок г99зэмембраны не отделяется от клетки и не нарушается его связь с цитоплазмой. Измерение на целой клетке при разрушении мембраны в кончике микропипетки (whole-cell) позволяет заменять ионный состав цитоплазмы и изучать на диализированных таким образом клетках ионные токи в режиме фиксации напряжения.
Ионные токи через небольшие мембранные фрагменты измеряют с помощью пипеток, у которых диаметр кончика соизмерим с размерами фрагментов. Сопротивление пипеток, заполненных раствором 150 ммоль/л KCl и погруженных в раствор такой же концентрации, приблизительно линейно зависит от площади отверстия кончика и варьирует от 1 до 5 МОм. Площадь отверстия кончика пипетки можно варьировать от 1 до 8 мкм2, изменяя степень нагрева спирали на последнем этапе вытягивания. Эти размеры находятся на грани разрешения светового микроскопа. Наружную поверхность покрывают гидрофобным материалом – силгардовой резиной. Особенностью незастывшего силгарда является его способность растекаться тонкой пленкой по поверхности стекла на несколько миллиметров, покрывая при этом и кончик микроэлектрода. Так как высокоомные контакты образуются только с чистым стеклом, эту пленку необходимо удалять только оплавлением микроэлектродов. При работе на мембранных фрагментах используется несколько типов пипеток.
Пипетки из тугоплавкого стекла получили в практике большее применение, чем пипетки из мягкого стекла. Первые имеют больше удельное сопротивление, чем мягкое стекло с более низкой температурой плавления. Вследствие этого вклад шума, обусловленного ёмкостью связи через стеклянную стенку в пипетках с тугоплавким стеклом меньше.
Пипетки из толстостенного тугоплавкого стекла имеют ряд преимуществ. Во-первых, при большей толщине стенок шунтирующая проводимость через стекло меньше. Во-вторых, на некоторых препаратах гигаомные контакты более стабильны и величина их образования значительно больше, чем для аналогичных тонкостенных пипеток Табл.1. Геометрические параметрыкончиков пипеток, изготавливаемых
из различных типов ст. капилляров
| Материал, из которого изготовлены пипетки | Площадь отверстиямкм2 | Площадь кольца, мкм2 | Ширина кольца, мкм | Угол конуса, град |
| Тонкостенные капилляры CEE BEE – мягкое стекло | 1.0 | 0,79 | 0,19 | 24 |
| Кимакс – твердое боросиликатное стекло | 1,2 | 0,82 | 0,2 | 20 |
| Алюминиевое – твердое алюмосиликатное стекло | 1,0 | 0,9 | 0,22 | 25 |
| Тонкостенные капилляры Jencons – твердое боросиликатное стекло | 1,01 | 1,71 | 0,39 | 10 |
При работе с некоторыми клетками, например, с изолированными с помощью ферментов клетками миокарда, гигаомные контакты иногда формируются спонтанно, при прикосновении кончиком пипетки к поверхности клетки высокоомный контакт формируется без подсасывания. В этом случае не наблюдается никаких деформаций клеточных мембраны, и площадь мембранного фрагмента можно оценить по значению сопротивления пипетки до образования контакта. Но в большинстве случаев контакт формируется только при небольшом разрежении, создаваемом внутри пипетки. При этом поверхность мембраны деформируется, часть её втягивается внутрь пипетки на 2-3 мкм, принимая Q-образную форму. Когда фрагмент изолируют механическим отведением кончика пипетки от поверхности клетки, его У-образная форма практически не изменяется. При дальнейшем подсасывании образуется сферический пузырёк. Если известна удельная ёмкость мембраны (как правило, она составляет 1 мкф/см2), площадь фрагмента можно оценить по величине его ёмкости; обычно площадь варьирует от 2 до 25 мкм2.
Электронные схемы для пэтч-кламп регистрации
Электронная схема пэтч-кламп регистрации должна иметь такие параметры, чтобы было возможным зарегистрировать передвижение всего лишь нескольких сотен элементарных электрических зарядов через малый участок клеточной мембраны. При максимально сниженных собственных шумах измерительной аппаратуры, токи через одиночные электровозбудимые Са-каналы, можно зарегистрировать только в нефизиологических условиях – например, при использовании растворов, содержащих Ва2+ вместо Са2+.
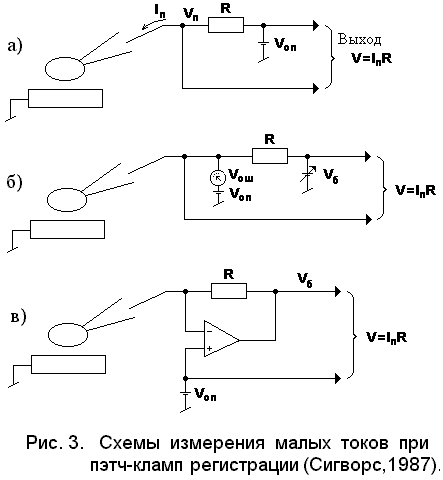
Стандартным методом измерения малых токов является регистрация создаваемого ими падения напряжения на высокоомном сопротивлении. На рис.3 представлен три электрические цепи, с помощью которых осуществляются подобные измерения. Наиболее удобно автоматическое поддержание требуемого значения Vб с помощью операционного усилителя, который представляет собой управляемый потенциалом источник напряжения ( рис.3, в).
Регистрация от целой клетки в условиях плотного контакта
Несмотря на то, что метод пэтч-кламп был разработан для регистрации токов через одиночные каналы, пэтч-пипетки можно с успехом использовать также для регистрации токов от целой клетки (РЦК), особенно когда размеры её невелики. После образования гигаомного контакта мембранный фрагмент под пипеткой можно разрушить, прикладывая к ней короткие импульсы отрицательного давления. Такая манипуляция в большинстве случаев не нарушает контакта пипетки с мембраной, и в результате между пипеткой и содержимым клетки устанавливается хорошо изолированный от омывающего раствора проводящий путь. Такой способ проникновения в клетку наносит ей гораздо меньше повреждений, чем введение стандартного микроэлектрода. Входное сопротивление небольших клеток велико по сравнению с эффективным сопротивлением кончика пипетки, поэтому при работе с ними электрические измерения можно проводить от участка мембраны намного большей площади, чем у исходного фрагмента. В отличие от разных вариантов ПК регистрации этот метод позволяет осуществить регистрацию от мембраны целой клетки, а не от маленького мембранного фрагмента.
Для регистрации на целой клетке пэтч-пипетки заполняют соответствующим раствором с низкой концентрацией ионов кальция. Пипетку прижимают к поверхности клеточной мембраны, в результате чего формируется высокоомный контакт. Затем потенциал пипетки изменяют до отрицательного значения и подают импульсы напряжения с амплитудой в несколько милливольт. На этом этапе производится компенсация быстрой ёмкостной компоненты, обусловленной ёмкостью держателя и стенок пипетки. К пипетке прикладывают импульсы отрицательного давления до тех пор, пока вновь не появятся ёмкостные переходные процессы (возникает дополнительный ток).
Регистрацию от целой клетки можно проводить в течение часа без визуальных или электрических признаков нарушения мембраны. Если исследуемая клетка плотно прикрепилась к культуральной чашке, плавным отведением её от пипетки можно получить конфигурацию outside-out. После такой процедуры клеточная мембрана быстро “зашивается” и клетка является интактной. Но содержимое клетки соответствует раствору в пипетке. Это используется в различных целях:
для изменения содержимого выбранной клетки с минимальным повреждением её мембраны.
Для проведения исследований с различными внутриклеточными растворами на одной и той же клетке путем смены регистрирующих пипеток.
Для регистрации в одном эксперименте интегральных токов и одиночных каналов.
Пипетки должны быть сравнительно широкими и иметь кончики с большим углом схождения.
Между раствором в пипетке и содержимым клетки происходит быстрый обмен, поэтому раствор для заполнения пипеток должен быть близок по своему составу к клеточному содержимому.
Таб. 2. Типичные электрические параметры,
получаемые при регистрации на хромафинных
клетках в конфигурации РЦК
| Последовательное сопротивление, Rs | 4 МОм |
| Ёмкость клетки, C | 5 пФ |
| Пост. Времени фиксации потенциала, RsC | 20 мкс |
| Пост. Времени обмена ионов, tоб | 5с |
| Сопротивление клетки, R | 10ГОм |
| Сопротивление утечки, Rут | 20ГОм |
| Шум (среднеквадр. значение, 0-400 Гц) | 0,15 пА |
Данные таблицы отличаются от значений, полученных с обычными стеклянными микроэлектродами по трем основным пунктам:
большинство клеток с диаметром менее 20 мкм при прокалывании обычным микроэлектродом повреждается, в то время как РЦК можно проводить даже на клетках диаметром менее 10 мкм, совершенно не нарушая их.
Сопротивление утечки микроэлектрод-клетка при использовании обычных микроэлектродов не превышает 500 МОм, в то время как соответствующее значение сопротивления утечки при РЦК могут быть 20 ГОм и более.
О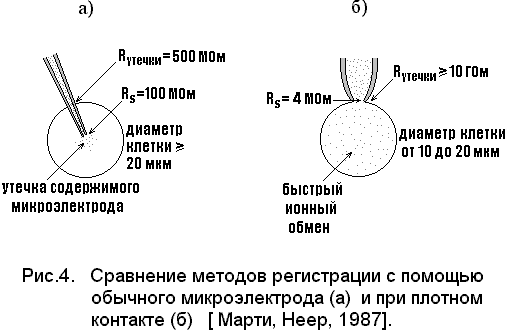
бычные микроэлектроды имеют сопротивление кончика Rs более 100 МОм, а типичные значения для РЦК на хромафинных клетках составляют 4 МОм.
Метод РЦК применяется для регистрации токов в мембране опухолевых клеток гипофиза, внутренних сегментов палочек сетчатки, островковых клеток поджелудочной железы, клеток миокарда, наружных сегментов палочек сетчатки, нейронов спинного мозга млекопитающих и др.
Пэтч-кламп регистрация при неплотном контакте
Метод регистрации от целой клетки в условиях плотного контакта позволяет регистрировать ионные токи, протекающие через небольшие мембранные фрагменты, которые содержат только один или несколько ионных каналов. Для получения информации о кинетике работы каналов или о плотности токов через мембрану проводится одновременная регистрация работы множественных каналов с использованием метода пэтч-кламп или применяется классический метод фиксации мембранного потенциала.
Регистрация при неплотном контакте по сути идентична регистрации одиночных каналов, за исключением того, что участок мембраны, на котором фиксируется потенциал, на 3 порядка больше, и при этом не требуется формирования гигаомных контактов.
Метод позволяет без труда получать стабильные электрофизиологические данные от локальных участков мембраны, используя интактные клетки практически без их изоляции или других видов обработки. Например, можно проводить измерения на целой мышце, не выделяя одиночные клетки. Отпадает необходимость и в использовании внутриклеточных микроэлектродов.
Однако, необходимо отметить, что наиболее корректную информацию о мембранной проводимости всё-таки даёт регистрация токов через одиночные каналы, так как позволяет избавиться от ряда артефактов, которые могут быть получены при регистрации макроскопического тока. Имеются, по крайней мере, четыре основные проблемы, с которыми приходится сталкиваться при регистрации макроскопических токов. Во-первых, трудно создать условия для регистрации тока только через каналы интересующего нас типа. Во-вторых, влияние последовательного сопротивления
может приводить к различию между истинным мембранным и поддерживаемым потенциалом. В-третьих, накопление ионов в примембранной области может вызывать зависящее от времени изменение тока через отдельные каналы. И, наконец, потенциал-зависимость проводимости открытого канала может быть ошибочно принята за потенциал-зависимость активации канала.
Литература:
Медведев С.С. Электрофизиология растений. – СПб.: Изд-во
С-Петебург. ун-та, 1998. – 184 с.
Регистрация одиночных каналов / под ред. Сакмана Б. и
Неера Э. – М.: “Мир”, 1991
Эккерт Р., Рэнделл.Д, Огастин Дж. – Физиология животных,
механизмы и адаптация. – М.: “Мир”,1991
Hedrich R., Schroeder.J.I. and Fernandez J.M. – Patch-clamp studies on higher plant cells: a perspective. // Trends biochemistry science. – 1987. – v.12. pp. 49-52&
Похожие работы
... является то, что рабочий стол 6 с обрабатываемыми образцами 5 размещается внутри данного устройства. Разрабатываемое оборудование позволит осуществлять имплантацию ионов азота с энергией 1 – 10 кэВ ( Дж) в металлы и сплавы, модифицируя их свойства в нужном направлении. Заключение Несмотря на большое количество исследований в области ионной имплантации, остаётся ещё множество вопросов, ...
... бы гораздо медленнее, чем в действительности. Причина этого в том, что общая проводимость мембраны в покое достаточно мала, и выход накопленных в результате потенциала действия положительных ионов из клетки занял бы несколько миллисекунд, а то и десятки миллисекунд. В действительности, возврат к уровню потенциала покоя происходит гораздо быстрее благодаря значительному увеличению другой мембранной ...
... ячейки под действием наложенного потенциала происходит электролиз – окисление или восстановление вещества. В кондуктометрическом анализе используют кондуктометрическую ячейку, в которой замеряют электрическую проводимость раствора. По способу применения электрохимические методы можно классифицировать на прямые, в которых концентрацию веществ измеряют по показанию прибора, и электрохимическое ...
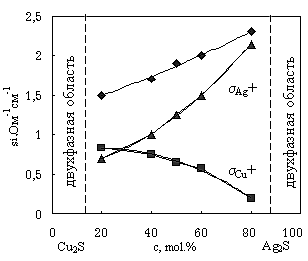
... с использованием сложных токовых электродов Ag/AgI. Как показано в [1], указанный метод, примененный к подобным системам с двумя сортами подвижных ионов, дает значение полной ионной проводимости. Разделение вкладов ионов меди и серебра в общую ионную проводимость твердых растворов (AgxCu1-x)2S было проведено комбинированным гравиметрическим и электрическим методом /1/ с использованием ячейки Ag/ ...
0 комментариев