Навигация
1. Тип рухів
відстань, які долають за одиницю часу
час амбуляції
частота підводжень на задні кінцівки
намагання уникати (стрибки)
латентний період
час без рухів
2. локомоція
“відвідування" певних відділів поля (центр або перефіричні кути)
Б) (Рухи) Переміщення частин тіла
маніпуляція об’єктами
принюхування
драпання
риття
оральний автоматизм
вмивання
вокалізація
візуальне обслідування
ІІ - Вегетативна нервова система
дефікація
урінація
ЧСС
ЧД
ІІІ - Надниркова активність
аскорбінова кислота наднирок
плазменні кортикостероїди
IV - Електроенцефалографія
гіпокамальний тета-ритм
електроміограмма
В основному це поведінкові ознаки, фізіологічні параметри, більшість з яких - варіанти рухової активності (амбуляція), завдяки легкості кількісних вимірів і явної надійності та вірогідності для інтерпретації. Вимірювання амбуляції звичайно робиться шляхом просторових підрахунків або вимірюванням пропорцій часу, який був використаний на мабуляцію. Виміри проводяться згідно:
1) характеристики їх чергових прагнень (цілі);
2) локальність;
3) латентність.
Амбуляція, яка була спрямована на втечу виміряна Ehrlich A. & Burns N. (1958). Місцезнаходження на периферії та в кутах (Morisson B.I. & Tratcher K. 1969), місцезнаходження біля стін (Vall F. D. 1970) - були використані як показники праці. Активність також може бути вивчена за її відсутності, за допомогою 2-х основних параметрів - латенція та фрізінг (“застивання, заморожування”). Латентний період вимірюється часом від початку експеримента до прояву певного типу поведінки. Звичайно, в якості латентного періода використовують час за який тварина залишає стартовий майданчик (Ivinskis A. 1968; W. Poley & I. P. Royce 1970). Теоретично може бути використана латентність будь - якого залежного параметра, але дослідники дають перевагу амбуляції.
Іншим обчислювальним параметром у “Відкритому полі” є відсутність активності - фрізінг. Він широко використовується як на індикатор стану стреса. Певно, це і є поведінкова реакція, яка на філогенетичній сходинці проявляється як сприйняття загрози (специфічна для кожного виду тварин). Факторами фрізінгу виступають раптові зміни, переміщення оточення (середові дислокації), фізичне обмеження рухомості та присутність хижаків. У “Відкритому полі” фрізінг - відповідь на подразники, які наведені вище і специфічні для кожного виду, іноді проявляються більше 1 години. Фрізінг може супроводжуватися тахіпноє та серцевою аритмією (M. A. Hofer, 1970). Другим варіантом довготривалої нерухомості виступає сон та уявний сон, який був досліджений Fox’ом & Spencer’ом І. М (1969). Тварини, у яких відмічали довготривалу нерухомість у “Відкритому полі”, швидко зупиняли рухи, посилювалася дефікація та урінація, вони навіть тремтіли при перенесенних в стартовий бокс. Уявний сон також відмічав Cummins R. A. у щурів, у яких виявлялося тремтіння, яке може бути результатом тахіпноє в стані нерухомості. M. A. Hofer (1970), Zynn R. (1966), відмічали уявний сон, як результат перезбудження, частота сну збільшувалась зі збільшенням часу досліду та часом між дослідами. Це скоріше звикання, яке вказує на те, що звикання до реакції збудження може супроводжуватися швидким станом дрімоти та сну. В цей період дрімоти може легко виникнути нереалізована орієнтовна реакція. Суб’єктивно, можливо диференціювати залякування від зовнішньої загрози та від псевдо-сплячки за характерними ознаками, такими як: широко відкриті очі, м’язова ригідність (скованість) проти постійного розвитку замкнутих очей та розслабленої пози, що супроводжують псевдосплячку. Ці дві форми поведінки наводять приклад реакції переляку та викликаної реакції збудження. Тестом для цих гіпотез може бути реакція реєстрації ЕЕГ тому, що орієнтовна реакція супроводжується гіпокамальним Q - ритмом, тоді як реакція переляку супроводжується десинхронізацією в гіпокампі. Отже, нерухомість у “Відкритому полі” може бути індикатором високої напруги (стресу).
Слідуючим критерієм активності є підводження - рерінг (Ivinskis A., 1968). В комплексі з амбуляцією він відображає стійкі індивідуальні ознаки “неспецифічний рівень збудженості”, який корелює з короткохвильовою активністю гіпокампу та з іншими поведінковими ознаками, також із варіантами інших поведінкових реакцій, в залежності від індивідуальних відмін в соматичних функціях, таких як, швидкість росту, температура тіла, тепловіддача, перевага певних видів їжі, ендокринні функції (Hat L. & Collova-Hemon 1968; Martinek Z. &Hat L. 1969). Сидіння виявляється при умовах, які зв’язані з низьким рівнем активності та негативно корелює з амбуляцією та рерінгом (Prescott R. 1970). Параметри, які не відносяться до рухів всього тіла, досліджені неповністю. Сюди відносяться: драпання, скрежет зубами, ЕМГ, серцеві ритми, ЕЕГ, які використовувалися для дослідження поведінки. Скрежет зубами було використано Hudhes’ом (1969) як можливий індикатор турботи (збентеженість), на тій основі, що воно може символізувати “поведінку заточення зубів з метою захисту”. Дихання та ЕМГ були використані Hofer’ом (1970) в дослідженні довготривалої амбуляції. Чищення шерсті (грумінг) і умивання, характеризують один і той же вид поведінки та визначають низьку вірогідність (Ivinskis A. 1968) та негативний зв’язок з показниками високоактивних станів (Prescott R. 1970).
Можливих параметрів вегетативної нервової системи багато, але на практиці вони звичайно лімітуються легко вимірювальними екскреторними та серцевими функціями. Характерним проявом пасивно-оборонної поведінки у щурів при переміщенні їх у нове незнайоме середовище, є вегетативна реакція у вигляді дефекації та урінації, а також зміненні рівня рухової активності. До форм пасивно - оборонної поведінки у тесті “Відкрите поле” наведені найменування: реакція дефікації і реакція урінації, що визначаються як “емоційність”, а рухову активність - як “амбуляцію”. Відображення пасивно-оборонної поведінки щурів через їх емоційність дослідив Hall (1934). У 1942 році Холл і Клайн порівняли 2-і лінії щурів за агресивністю. Вони з’ясували, що неемоційні щури були більш агресивні ніж емоційні. В зв’язку з їх дослідами можливо провести деяку аналогію між емоційністю і пасивно-оборонною поведінкою, як між неемоційністю та активно-оборонною поведінкою. Подальший розвиток методика “Відкритого поля” отримала у працях Broadhurstа (1957), який застосував не лише яскраве світло, але і гучний звук. Він виявив, що коефіцієнт надійності цього тесту дорівнював 0,82. таким чином, Broadhurst і Holl провели вдалий експеримент та вивели 2 лінії щурів: емоційну, за їх термінологією “реактивну” (MR) та неемоційну, або “нереактивну” (MNR). Ці дві лінії щурів вірогідно відрізняються за багатьма морфологічними та функціональними ознаками (Broadhurst & Eysenck, 1964), а також було відзначено, що дефікація і рухова активність (амбуляція) мають полігенну природу успадкування (Broadhurst P. L. 1959) [14].
Як прояв вегетативної реакції дефікація залишається домінуючим індексом емоційності, значення якої було підтверджено факторним аналізом (Whimdey & Denenberg, 1967). Крім дефікації враховується урінація за частотою прояву, присутністю або відсутністю, кількості або якості елімітуючої суміші з дефікацією (Tobach, 1966).
3. Методи та матеріали дослідження. 3.1 Методика вироблення штучного циркадного біоритму у самців щурів лінії Вістар
Дослідження проводилося на 60 самцях щурів лінії Вістар віком 7-7,5 місяців, масою 290-420 г, довжиною 330-410 мм, тривалість життя 2,5-3,5 роки, іноді до 5-6 років. Статева зрілість - 1,5-2 місяці.
За характером дії та інтенсивністю освітлення на початку експеримента було сформовано 3 групи:
К - контрольна (інтактна) - тварини знаходились при дії природного освітлення;
Д-Н - при дії штучного освітлення на протязі 12 годин на добу: з 6 години ранку до 18 години вечора);
Д - при дії цілодобового штучного освітлення.
Експеримент тривав 5 місяців. Кожна група утримувалася в окремих приміщеннях. Тварини знаходилися в стандартних клітках по 5 самців в кожній, при харчуванні ad libitum, вживання води було вільне.
Для створення штучного освітлення використовувалися лампи накалювання потужністю 100 Вт. Кожна клітка з тваринами мала свою власну лампу, яка була розміщена від неї на відстані 0,5 м. Рівень освітлення в приміщеннях зі штучним освітленням складав 300 лк. [8]
Саме така інтенсивність освітлення дає можливість ліквідувати розбіжність між циркадними ритмами у тварин. Якщо використовувати її нижчі або вищі показники, це призведе до суттєвого зрушення ритму виділення мелатоніну і, таким чином, до розбіжності ритмів організму в цілому. [10]
3.2 Методика проведення тесту “Відкрите поле”
Методика “відкритого поля” проводилась з використанням устрою, який являє собою квадрат розміром 1м х 1м, обмежений бортами висотою 40см. Вся площа поля розкреслена лініями на 25 квадратів зі стороною 20 см. [14] Над центром поля на відстані 0,5 м розташована лампа накалювання потужністю 100 Вт. Устрій знаходився у затемненому приміщенні. Перед розміщенням у “відкрите поле” щурів поодинці саджали у коробку і через 10 хв проводили тестування.
Спостереження за тваринами відбувалися на протязі 3х днів, в один і той же час доби за однакових умов освітлення і температури, за відсутністю сторонніх запахів і шуму. Час експозиції кожної тварини у “відкритому полі” становив 5 хв. Виборка тварин з кожної групи становила по 10 самців.
В індивідуальній поведінці тварин реєстрували їх горизонтальну рухову активність (амбуляції, локомоції); вертикальну активність (рерінг, підведення на задні лапки); латентний період (латенція); дефікацію (кількість болюсів - фекальних шариків); грумінг (кількість вмивань) та урінацію. [12,13]
4. Поведінкові реакції щурів які утримувались у різних вітлових режимах 4.1 Поведінкові реакції самців щурів контрольної групи
1) У самців даної групи був відсутній латентний період. Взагалі, латентність характеризується часом від початку досліду до появи певного типу поведінки і стартової позиції (Ivinskis A., 1968, Poley W. &Royce I. P., 1970). У нормі латентний період збільшується на 3 день (Furechtgott E. at al., 1961), злегка знижується на 2 день (Ader R., 1968). В даному випадку відсутність лацентації самців контрольної групи свідчить про їх високу активність і швидку адаптацію до умов навколишнього середовища.
2) Амбуляція щурів цієї групи протягом експерементальних днів поступово зменшувалась з 7,68 до 2,82. (мал.1) (Амбуляція - локомоція - це кількість перетнутих квадратів твариною всіма чотирма лапами; це факторний комплексний параметр, що відображає дослідницьку діяльність і емоційність). Це говорить про наявність у них редукції страху. Отримані дані відповідають літературним. Адже, за Whimbey A. E., Denenberg’ом (1969) частота амбуляції через інтервал часу змінюється від позитивної - в 1й день, до негативної на 2й день і т.д. малюнок 1
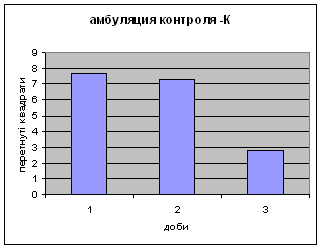
3) Вегетативні показники.
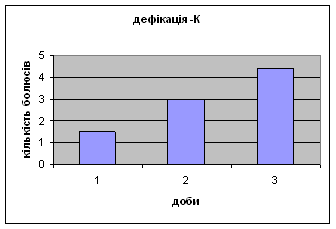
Дефікація зростала з 1,5 до 4,4 (мал.2), що відповідає літературним даним. Так, за Broadhurst’ом Р.Н. (1969) виявлено тенденцію до її зменшення протягом 3х днів спостереження. Загалом, дефікація та урінація - це перші показники емоційності тварин. Таким чином, отримані дані свідчать про те, що у самців цієї групи знижується емоційний стан і спостерігається розвиток звикання до умов. Відсутність латентного періоду та позитивна кореляція між амбуляцією та дефікацією, також свідчить на цю користь.
Малюнок 2
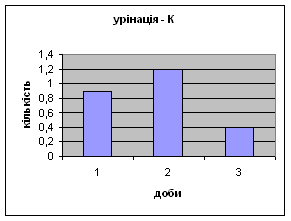
Спостерігалось зменшення показників урінації у самцув контрольної групи від 0,9 до 0,4 (мал.3), хоча на другий день експеременту спостерігалось деяке підвищення сечовипускання що. Отримані дані а також позитивна кореляція власне цього показника з амбуляцією, підтверджує думку про редукцію страху у цих щурів.
Малюнок 3
4) Протягом експерименту у піддослідних тварин зменшувались показники рерінгу (вставання на задні лапки, це основний показник активності тварин і використовується він як індекс активності) принюхування (мал.4 і мал.5), та зростали показники грумінгу (вмивання) (мал.6), що разом з описаними вище даними говорить про те, що у самців даної групи знижується емоційність і розвивається звикання до стресових факторів навколишнього середовища.
Малюнок 4 Малюнок 5
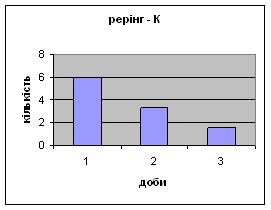
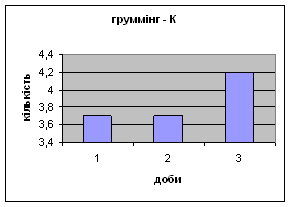
Малюнок 6
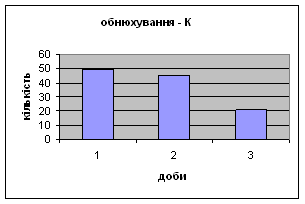
4.2 Поведінкові реакції самців щурів Д-Н групи
1) Відсутня лацентація.
43 2) Амбуляція у самців Д-Н-групи протягом 3х днів зменьшувалась з 16,2 до 7,11. (мал.7) Тобто спостерігається більш поступове зниження цих показників в порівнянні з контрольною групою, і, в той же час, більша рухова активність, що проявлялась в перетнутих квадратах.
Малюнок 7
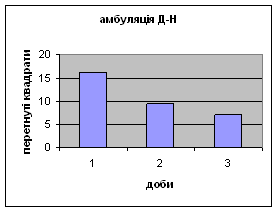
3) Вегетативні показники.
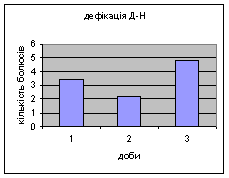
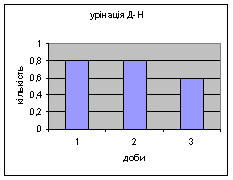
Показники дефікації, в порівнянні з іншими групами, мали свою криву: на другий день експерименту кількість болюсів різко зменшилась з 3,4 до 2,2 (мал.8), а потім почала поступово зростати до 4,8. Ці дані свідчать про те, що у самців Д-Н-групи відбувається більш повільніша редукція страху і емоційності, в порівнянні з іншими групами піддослідних тварин. Позитивна кореляція між амбуляцією і дефікацією також дає підстави вважати те, що існує зниження тривоги у даних самців і загальна їх адаптація до умов. Урінація протягом перших 2х днів експерименту була незмінна - 0.8, а далі почала знижуватись до 0,6 (мал.9).
Малюнок 8 Малюнок 9
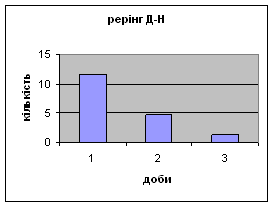
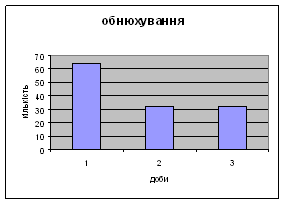
4) Також специфічність самців Д-Н-групи проявилась і при аналізі показників принюхуванні та рерінгу. Загальне число принюхувань, в порівнянні з іншими групами набагато вище, так в перший день дослідження зареєстровано 64,4 принюхувань. Як і в контрольній групі, даний показник зменшувався (до 32,3) (мал.11). Найбільший коефіцієнт активності (рерінг) був зареєстрований саме в цій групі тварин.
Протягом перших двох днів досліду він майже був незмінний - 11,7, а потім різко почав знижуватись аж до 1,3 (мал.10). Все вище описане говорить про те, що дані самці Д-Н-групи більш важче пристосовуються до умов середовища, особливо в перші дні, де адаптація майже не спостерігається. Але потім, ці піддослідні тварини активно наздоганяють в цьому аспекті тварин інших груп. Про що свідчить позитивна кореляція дефікації, амбуляції та урінації між собою, а також зменшення показників рерінгу та принюхування. Таким чином, у самців Д-Н-групи також відбувається редукція емоційності, тривоги, тобто розвивається звикання до стресових факторів.
Малюнок 11 Малюнок 10
4.3 Поведінкові реакції самців щурів Д-групи
1) Відсутня лацентація.
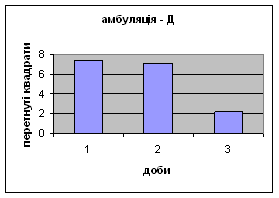
2) У самців Д-групи спостерігалась найнижча локомоція в порівнянні з тваринами інших груп. Це говорить про те, що світло для них був звичним фактором існування. На відміну від інших груп, серед переміщення даних тварин по “полю" було зареєстровано перетинання майже всіма піддослідними центра “поля”. Такий характер амбуляції не спостерігався серед інших експериментальних груп. Деякі Д-самці навіть тривалий час знаходились безпосередньо під лампою. Таким чином, на основі отриманих фактів, можна сказати, що тварини цієї групи майже не відчували страху бо дія світла для них була звичною. Показники амбуляції протягом 3х днів знижувалась з 7,38 до 2,19 (мал.12).
Малюнок 12
3) Вегетативні показники.
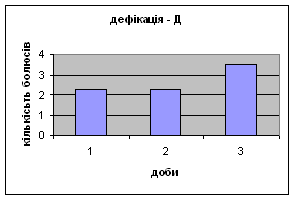
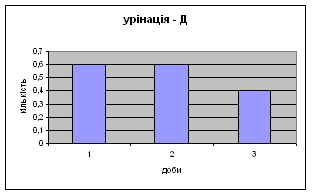
Показники дефікації зростали з 2,3 до 3,5 (мал.13), що говорить про швидке звикання до умов існування. Позитивна кореляція між показниками дефікації та локомоції свідчать також на цю користь. Найменша кількість урінацій (від0,6 до 0,4) (мал.14) була зареєстрована саме в цій групі самців, яка також мала позитивну кореляцію з амбуляцією.
Малюнок 12 Малюнок 13
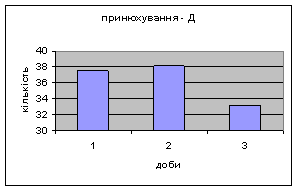
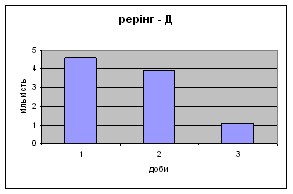
4) Самці Д-групи за показниками рерінгу були найменш активними серед експериментальних груп. Як і в попередніх групах дані показники цього параметру зменшувались (з 4,6 до 1,1) (мал.15). Але в порівнянні з К і Д-Н - групами, самці Д-групи протягом перших 3х днів експерименту проявляли стійки в напрямку до лампи, що не спостерігалось серед інших груп. Знову, це підтверджує думку про те, що дані тварини не відчували страху перед дією світла. Це, а також незначне зменшення кількості принюхувань (від 37,5 до 33,2) (мал14), дає можливість зробити висновок про швидкий розвиток у самців Д-групи звикання до стресових факторів середовища.
Малюнок 14 ёМалюнок 15
Висновки
0 комментариев