Ю.А. ЛАБАС, к.б.н., вед. научн. сотр., А.В. ГОРДЕЕВА, аспирант, Ин-т биохимии им. А.Н. Баха РАН
На протяжении полутора веков, прошедших после первого издания «Происхождения видов» Ч.Дарвина (1859), триада факторов эволюции «наследственность – изменчивость – естественный отбор» постоянно подвергалась нападкам. Открытие законов Г.Менделя, достижения популяционной и молекулярной генетики внесли, конечно, серьезные коррективы в первоначальные представления о процессе эволюции, но стержневая идея отбора уцелела.
Одна из не решенных теорией эволюции проблем – появление в филогенезе новых признаков. Считается, что они возникают при замене одних биологических функций другими, например парные плавники рыб заменились конечностями четвероногих наземных позвоночных, а затем передняя пара конечностей – крыльями, которые появились у птерозавров, птиц и рукокрылых млекопитающих.
В то же время известно, что новые функции часто представляют собой результат закрепления и дальнейшего усовершенствования отбором случайных макромутаций – скачкообразных изменений генетической информации, приводящих к обретению организмом новых наследственных признаков, которые дают их обладателям те или иные преимущества в сравнении с исходным генотипом. В результате такие «удачные находки» закрепляются естественным отбором.
Ясно, что у организма уже должны существовать необходимые предпосылки, чтобы за счет одной макромутации или их небольшого числа возникла новая функция. Чем ближе к истокам жизни в эволюции появлялись новые биологические функции, тем труднее объяснить, как это происходило. Так, до сих пор неизвестно, когда и как в клетках «обосновались» рибосомы. Столь же загадочно происхождение клеточных мембран, самой генетической информации и т.д. и т.п.
В «Происхождении видов» Дарвина есть специальная глава «Частные трудности теории естественного отбора». В ней рассматриваются в основном два примера – возникновение органов свечения у разных организмов и электрических органов у рыб. Дарвин не смог объяснить появление этих органов, исходя из своей теории, но ему было ясно: их функции связаны с поведением животных. Мишенью отбора могли быть лишь такие признаки неопределенной изменчивости, при наличии которых у отдельных особей несветящихся организмов вдруг возникало свечение, хорошо заметное в темноте, или когда у неэлектрических рыб внезапно рождались индивиды, генерирующие вполне ощутимые электрические разряды.
Как ни странно, со времен Дарвина происхождению биолюминесцентных и электрических органов было посвящено сравнительно мало работ. За полтора столетия не сформировалось общепризнанного мнения о том, как могли возникнуть эти феномены. Мы предлагаем читателю свою точку зрения, но только в отношении биолюминесценции.
Светятся многие и по-разному
Биолюминесценцией называют свечение живых организмов, хорошо видимое в темноте.
Сейчас известны многие сотни биолюминесцентных видов, даже родов насчитывается более 700. Светиться способны бактерии, одноклеточные эукариотические организмы (жгутиконосные водоросли динофлагелляты, радиолярии), грибы и подвижные многоклеточные животные разных типов – от полипов, медуз и гребневиков до кальмаров, ракообразных и рыб. Большинство светящихся существ морские, среди них много глубоководных. Из наземных организмов светятся отдельные виды грибов, земляных червей, улиток, многоножек, комаров и жуков. Пресноводных биолюминесцентных видов пока обнаружено чрезвычайно мало: новозеландский брюхоногий легочный моллюск Latia neritoides и несколько видов паразитических бактерий.
У многоклеточных животных обычно излучает свет не все тело, а только специальные клетки фотоциты, часто сгруппированные в особые органы свечения – фотофоры. У некоторых рыб и кальмаров они очень сложно устроены (отражатель, линза и т.д.). Иногда фотофоры светятся непрерывно благодаря обитающим в них симбиотическим фотобактериям. Но обычно свечение импульсное, контролируемое нервной системой. Оно бывает внутриклеточным или секреторного типа. В последнем случае светящееся вещество (вернее, смесь продуктов синхронной секреции двух разных желез) извергается из организма в виде покрывающей его слизи или расплывается вокруг него в воде, как большое световое облако.
Субстраты и ферменты
В 1885 г. французский ученый и врач Р.Дюбуа доказал, что светящееся вещество биолюминесцентного организма состоит из двух компонентов и только один из них устойчив к нагреванию до 60–100 °С. Позже выяснилось, что термостойкое низкомолекулярное вещество, названное люциферином, представляет собой субстрат биолюминесцентной реакции, которую катализирует разрушающийся от нагревания фермент люцифераза.
Названия эти условные, у разных организмов такие субстраты и ферменты совершенно несхожи. В дальнейшем оказалось, что у некоторых организмов за свечение ответственны фотопротеины – стойкие комплексы люциферина и особого белка, который обычно «по совместительству» выполняет функции фермента люциферазы. У грибов люциферазы вообще нет, люциферин светится без нее, реагируя с одной из активных форм кислорода – супероксидом (рис. 1).
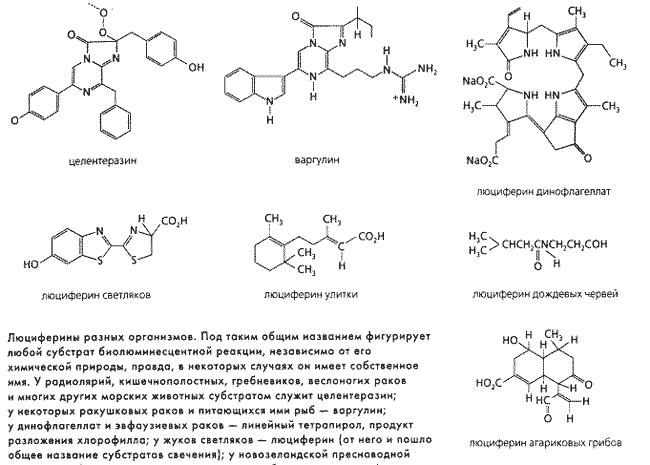
Рис 1.
Удивительно, что далеко не все светящиеся животные сами синтезируют свои люциферины. Особого внимания заслуживает в этой связи целентеразин (от Coelenterata – кишечнополостные), производное имидазолпиразина. Он выполняет функцию люциферина не только у кишечнополостных, но также у радиолярий, гребневиков, щетинкочелюстных червей, некоторых рыб и кальмаров, креветок и, наконец, веслоногих и ракушковых планктонных рачков. Возможно, многие из перечисленных животных не способны сами синтезировать целентеразин, а получают его, питаясь этими рачками. Такое предположение высказал в 2001 г. американский ученый С.Хэддок, когда обнаружил, что медуза экворея утрачивает способность к биолюминесценции, если в ее рацион не входят светящиеся веслоногие рачки.
Уже ранее было известно, что так же обстоит дело с рыбами Porychthis и эвфаузиевыми раками Meganyctiphanes. Первые получают люциферин от ракушковых рачков рода Vargula (у них это варгулин – гетероциклическое соединение, несколько отличающееся от целентеразина), вторые – от динофлагеллят, у которых роль люциферина выполняет линейный тетрапирол, продукт разложения хлорофилла.
По мнению Л.Тиси и Дж.Мюррея, высказанному ими совсем недавно на Международном симпозиуме по био- и хемилюминесценции, светляки тоже люциферин не синтезируют, а заимствуют у бактерий.
Между тем совсем недавно в Каспийском море, где никогда не было никаких светящихся организмов, в том числе и планктонных рачков, вдруг появился новосел – гребневик Mnemiopsis leidui, уроженец Атлантики. Оказалось, что мнемиопсис и в Каспии продолжает светиться «назло Хэддоку» и, стало быть, сам синтезирует целентеразин, а не заимствует его у рачков. Или, может быть, небиолюминесцентные каспийские рачки, которыми он питается, целентеразин все-таки синтезируют, но используют по какому-то иному назначению, например, в качестве антиоксиданта?
Вполне очевидно, что не могут питаться рачками и одноклеточные морские организмы радиолярии. Тем не менее и у них есть целентеразин. Как объяснить такие факты в свете данных Хэддока? Поступлением этого вещества с небиолюминесцентной пищей? Независимой эволюцией? «Кражей» генов, ответственных за синтез целентеразина? Пока эти вопросы остаются без ответа.
Приспособительный смысл свечения
Биолюминесцентный свет «холодный». Он не порождается высокой температурой источника, в отличие, например, от света обычной лампы накаливания. Тем не менее вряд ли какой-нибудь организм может позволить себе роскошь светиться без надобности. Энергетически это слишком дорогое удовольствие. Немногие из светящихся организмов (колонии бактерий, высшие грибы, многоножки, некоторые насекомые и др.) испускают свет непрерывно (статически), чем в темноте привлекают зрячих животных. Такой свет, как полагают, способствует попаданию светящихся паразитических бактерий в организм нового хозяина и распространению грибными комариками спор светящихся грибов. Мелких насекомых привлекает статическое свечение насекомоядных личинок новозеландских комаров Arachnocampa – они собственным телом подсвечивают свою ловчую сеть.
Между тем огромное большинство биолюминесцентных существ, включая медузу экворею, генерирует более или менее короткие (0, 1–1 с) световые вспышки в ответ на внешние механические и другие раздражения. Чаще всего такой свет дезориентирует зрячих хищников или отпугивает быстро движущихся крупных животных, способных повредить желеобразный светящийся организм (медузу, древовидную колонию полипов, гребневика) при случайном столкновении с ним. Поэтому защитная импульсная биолюминесценция обычно возникает синхронно с двигательной реакцией испуга. У гребневиков Bolinopsis, колониальных оболочников Pyrosoma, эвфаузиевых раков Meganyctiphanes такая реакция механически потревоженной особи вызывает подражательное свечение других особей, находящихся поблизости.
В прочих случаях статическое или импульсное свечение организмы используют для внутривидовой коммуникации, в том числе как сигнал, привлекающий особей другого пола. Самки американского светляка Photuris versicolor вначале подманивают специфичной «световой морзянкой» самцов своего вида и спариваются с ними, а затем начинают генерировать «морзянку» для самцов чужого вида, чтобы закусить ими.
У глубоководных рыб удильщиков имеется надо ртом подвижный отросток – «удилище», – на кончике которого находится световая приманка для жертвы. Рыбы Malacosteus niger освещают ближнее пространство красным светом, более никому из глубоководных животных не видимым, и тем спасаются от врагов. Рыбы Leognathus equulus используют свечение своего брюха для маскировки на светлом фоне водной поверхности и т.д.
Всего насчитывается больше 30 биохимических вариантов биолюминесценции, возникших в эволюции независимо и в ряде случаев, видимо, недавно. О недавнем происхождении отдельных вариантов свидетельствует видовая «чересполосица», т.е. близкое родство светящихся и небиолюминесцентных видов. Так, те и другие присутствуют в одном и том же роде у планктонных веслоногих рачков Oithona и Pseudocalanus или у колониальных гидроидных полипов Obelia. У агариковых грибов Pannelus stipticus, встречающихся в Европе и Северной Америке, плодовые тела ярко светятся преимущественно у американских штаммов. У этих грибов, биолюминесцентных фотобактерий и водорослей динофлагеллят часто возникают небиолюминесцентные мутанты. Сама же биохимическая природа свечения нередко существенно различается даже у близких родичей, например у разных видов веслоногих, а также ракушковых рачков.
Как возникли биолюминесцентные системы?
Итак, Дарвин отметил, что появление биолюминесценции трудно объяснить с позиции теории естественного отбора. Полезный эффект свечения всецело связан со зрительным восприятием и ответным поведением животных. Следовательно, повышать шансы выживания может только биолюминесценция, хорошо заметная в темноте. Кроме того, световые вспышки должны быть приурочены ко вполне определенным ситуациям, скажем, сопровождать защитную двигательную реакцию, вызванную приближением хищника, и отпугивать или дезориентировать его. Иначе свечение не даст никакой пользы, скорее нанесет вред.
Плавный, постепенный переход к свечению, хорошо заметному адаптированным к темноте глазом, невозможен – ведь ниже порога видимости не будет влияния на отбор. Следовательно, у небиолюминесцентных видов должны время от времени возникать мутанты, ярко светящиеся, например, при испуге, что повышает шансы спастись от хищника. Только так может начинаться процесс естественного отбора, приводящий к дальнейшему усовершенствованию биолюминесцентной системы.
Заметим, пока таких мутантов – «белых ворон» среди заведомо небиолюминесцентных видов – никто не видел, не искал, не пытался получить действием мутагенов. Описаны только упомянутые уже штаммы гриба P. stipticus, несветящиеся мутанты фотобактерий и водорослей динофлагеллят.
Кислород и его активные формы
Что же служит непосредственной причиной свечения? Показано, что для свечения всегда необходим молекулярный кислород или его активные формы. Атом кислорода, помимо основного, устойчивого триплетного состояния, может находиться в нескольких нестабильных, или возбужденных, состояниях. Одно из них – синглетное. Возврат электронов из синглетного состояния в триплетное сопровождается испусканием фотонов, правда, «маленьких», инфракрасных. Свечение же организмов обычно синее или зеленое.
Причина, как полагают, в том, что из возбужденного состояния в основное переходят сразу два или более атомов кислорода и энергии этих переходов суммируются. Одновременно рвутся О–О-связи в циклической и очень нестойкой диоксетановой перекиси, в которую превращается субстрат люциферин при окислении молекулярным кислородом или его активными формами. Их несколько, но главных три: супероксид, перекись водорода (H2O2) и чрезвычайно агрессивный окислитель гидроксил-радикал (OH–).
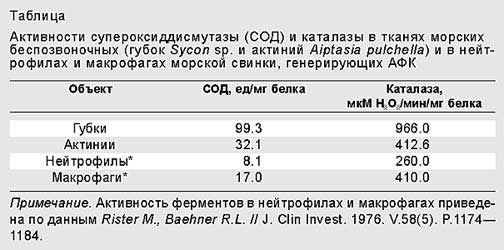
Активные формы кислорода (АФК) играют огромную роль в жизни организмов. С одной стороны, эти формы – «полуфабрикат» и «брак» дыхания митохондрий – способны окислять что попало в живом организме: ДНК, РНК, белки, липиды. Для защиты от этой опасности организмы вынуждены постоянно синтезировать или потреблять с пищей разнообразные антиоксиданты (в их числе витамины А, С, Е,
-каротин и др.), а также использовать такие ферменты, как супероксиддисмутаза, преобразующая О2– в перекись водорода H2O2, и каталаза, ускоряющая образование воды из перекиси.
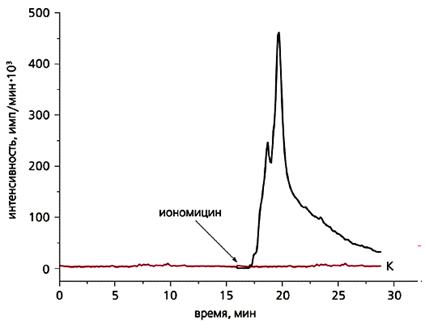
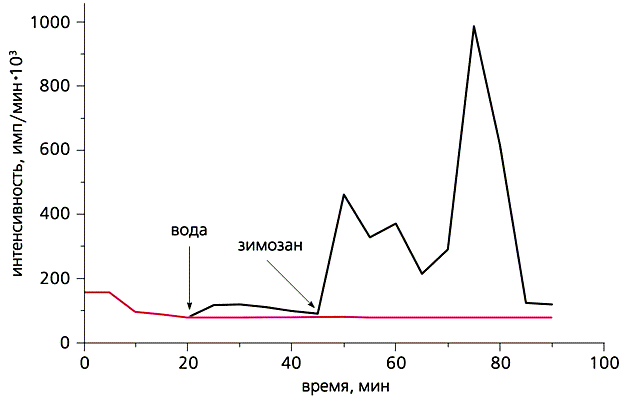
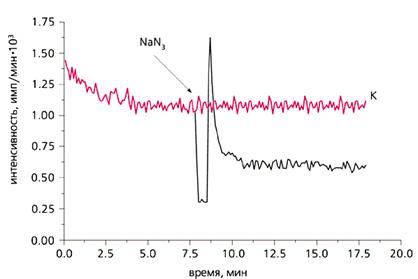
С другой стороны, в умеренных количествах АФК продуцируются в любом живом аэробном организме благодаря специальным ферментам, не связанным с процессом дыхания в митохондриях. АФК в низкой концентрации необходимы во многих жизненно важных процессах. Так, эти формы секретируются в импульсном режиме фагоцитами и некоторыми другими клетками для уничтожения паразитических микроорганизмов. АФК участвуют в регуляции клеточного деления, оплодотворения яйцеклетки, а также в запуске «запрограммированной смерти» клеток (апоптоза), в управлении тонусом кровеносных сосудов и т.д. Недавно мы обнаружили, что клетки поверхности ряда водных многоклеточных животных, подобно фагоцитам, тоже выделяют АФК, вероятно, для защиты от гнилостной микрофлоры. Природными (абиогенными) источниками АФК служат ионизирующая радиация, фото- и электрохимические реакции в воде. В частности, АФК образуются при фотолизе воды под действием УФ-излучения.
Мы затронули эти сложные вопросы потому, что работы последних лет доказывают: практически любой люциферин – высокоэффективный антиоксидант, а некоторые люциферазы – пероксидазы. Ферменты такого типа могут дезактивировать H2O2, окисляя с ее помощью определенные субстраты.
Таким образом, вполне возможно, что субстраты и ферменты биолюминесцентных реакций выполняют функции антиоксидантов у ближайших несветящихся родичей биолюминесцентных организмов. Поэтому достаточно одной малой наследственной «поломки» исходной (антиоксидантной) реакции, чтобы в ходе ее появилось хорошо заметное свечение и возник новый признак – биолюминесценция. Его может закрепить естественный отбор.
Как уже говорилось, природой таких предполагаемых нами «излучающих» мутаций пока, к сожалению, никто специально не занимался. Само их обнаружение – дело будущего.
Единого мнения о том, какие именно «добиолюминесцентные» функции выполняли молекулы люциферина и люциферазы, у ученых нет.
Одно время думали, что люциферазы произошли от ферментов, которые некогда, на заре жизни, защищали организмы от молекулярного кислорода, появившегося в атмосфере в больших количествах в связи с фотосинтезом. Позже, однако, выяснилось, что эти ферменты у разных групп организмов не сходны между собою. В эволюционном отношении они, видимо, гораздо моложе, чем сравнительно менее разнообразные люциферины, их субстраты. Да и биолюминесценция обретена многими организмами, как уже говорилось, не так уж давно и независимыми путями.
В 1998 г. группа ученых во главе с Ж.Р. Рийзом (Бельгия) предложила гипотезу, согласно которой «добиолюминесцентная» функция люциферинов состояла в защите морских организмов от АФК, образующихся в верхних слоях океана под действием ультрафиолета. Если такой организм переселялся на большую глубину, антиоксиданты там, в темноте, ему становились не нужны, и их можно было использовать для свечения.
Другие авторы полагают, что люциферины могли предохранять организм от собственных АФК – «неизбежного зла» при аэробном дыхании. Как известно, далеко не весь кислород восстанавливается в основной дыхательной цепи митохондрий. Нет-нет да образуются АФК и начинают «бесчинствовать» в клетках. Чем больше потребляется кислорода, тем больше этого «брака». От него-то, считают, и спасали «пробиолюминесцентные» системы.
По третьей версии, люциферины в большинстве своем – «вещества-чужаки» (ксенобиотики), попадающие в организм с пищей, а перекисное окисление уменьшает их токсичность.
Наконец и наша точка зрения. Один из авторов этой статьи, Ю.А. Лабас, еще в 1990 г. на Х конференции, посвященной памяти Л.А. Орбели, обратил внимание на то, что биолюминесцентными бывают главным образом клетки, по тем или иным причинам сами секретирующие АФК в повышенных количествах с помощью специальных ферментов, которые не связаны с митохондриями. Так, у некоторых дождевых червей и колониальных асцидий, если их потревожить, начинают ярко светиться фагоциты, выходящие на поверхность тела вместе с целомической жидкостью и разрушающиеся там (у асцидий – под прозрачной мантией). Напомним, что одна из функций фагоцитов – импульсная секреция АФК для уничтожения бактерий.
У большинства других многоклеточных животных ярко светятся фотоциты, расположенные на поверхности тел или, как у кишечнополостных и гребневиков, – в энтодерме.
Эпителиальные клетки, которые выделяют в воду светящиеся вещества, сами способны производить АФК в импульсном режиме, видимо, в качестве антимикробного средства. Это только что подтвердили наши эксперименты.
«Рекордсмены» в отравлении окружающей среды активными формами кислорода и особым ядом сакситоксином – морские планктонные динофлагелляты, в том числе светящиеся, чрезмерное размножение которых является причиной так называемых красных приливов, губительных для всего живого, кроме самих водорослей.
Не исключено, что и фотобактерии, паразиты и симбионты, выбрасывают АФК в окружающую среду. У ряда видов паразитических небиолюминесцентных бактерий такая секреция уже доказана, а у фотобактерий ее пока просто не искали. Паразитические бактерии, выделяя АФК, вероятно, разрушают таким способом ткани «хозяина», а, возможно, еще и уничтожают конкурентов.
Таким образом светятся (постоянно или в импульсном режиме) в основном клетки, которые сами секретируют АФК для той или иной цели, главным образом для защиты. Поэтому мы полагаем, что биолюминесценция – это результат наследственных изменений механизмов, защищающих подобные клетки от самоуничтожения.
Характерно, что многие выделяемые в импульсном режиме люциферины или их комплексы с белками, т.е. фотопротеины, светятся даже без люциферазы, реагируя не с молекулярным кислородом, а с АФК. Подобным образом излучают свет в эксперименте люциферин ракушковых рачков рода Vargula и фотопротеин двустворчатых моллюсков рода Pholas (оба эти соединения – варгулин и фолазин – используют как сверхчувствительные индикаторы АФК).
Выделение светящихся соединений во внешнюю среду водными организмами напоминает защитный прием наземного жука бомбардира (род Brachinus). Потревоженный, он довольно метко стреляет во врагов жгучей смесью трех веществ: восстановленного субстрата (гидрохинона), фермента (пероксидазы) и перекиси водорода. Мы не исключаем, что в состав излучающих веществ, выбрасываемых водными животными, первоначально тоже входила (или даже входит, это не проверено) в качестве третьего компонента перекись водорода или другие АФК и что светящаяся смесь происходит от жгучей защитной смеси.
Есть много примеров, подтверждающих связь биолюминесценции с активными формами кислорода. Приведем некоторые.
У многощетинковых червей из семейства Polynoidae внутриклеточные световые вспышки вызываются образованием супероксида в своеобразных клеточных органоидах – люмисомах. Продуцируется он специальным ферментом, активируемым повышенной цитоплазматической концентрацией ионов кальция. А светится от супероксида другой, находящийся в тех же клетках белок, полиноидин.
У динофлагеллят и морских перьев рода Renilla люциферин не может реагировать с люциферазой и кислородом, пока остается связанным с особым белком. Когда клетка возбуждена, в ней повышается концентрация Ca2+, что влечет за собой высвобождение люциферина. Только после этого он может реагировать с кислородом и люциферазой.
Наконец, у радиолярий, гидроидных полипов и гребневиков белок, который связан с люциферином, обрел функцию люциферазы, вероятно, вторично в процессе эволюции. Достаточно лишь повысить концентрацию Ca2+ в клетке – и перекись люциферина, будучи в комплексе с таким белком, начинает светиться при распаде. С эволюционной точки зрения немаловажно, что кроме ионов кальция свечение подобных фотопротеинов могут вызывать активные формы кислорода.
Итак, у разных организмов импульсное свечение обеспечивается отнюдь не одинаково. У асцидий, многощетинковых и дождевых червей оно связано с импульсной генерацией самих АФК. У динофлагеллят и морских перьев – с порционным выбросом люциферина – антиоксиданта, защищающего организм от повреждений, вызываемых этими агрессивными окислителями. У светляков вспышки создаются, как полагают, сложным механизмом импульсной подачи кислорода к месту внутриклеточной локализации люциферина и люциферазы. Мы здесь рассматривать этот механизм не будем. Важнее попытаться ответить на вопрос: как у несветящихся родителей может вдруг появиться более или менее ярко светящийся потомок?
Данных пока нет, поэтому остается лишь предполагать. В одних случаях может изменяться реакция синтеза того или иного антиоксиданта и в результате – его структура. Обновленный антиоксидант начинает ярко светиться, реагируя с АФК. В других случаях антиоксидант (свой или полученный с пищей) в результате мутации попадает в клетки, где присутствует фермент – будущая люцифераза, и тогда яркое свечение в реакции с молекулярным кислородом или перекисью водорода обеспечено. Если же мутация меняет структуру фермента, с участием которого окисляется восстановленный субстрат, такой процесс тоже может сопровождаться свечением.
Это наши соображения по поводу возникновения биолюминесценции – эволюционного события, которое великий Дарвин затруднялся объяснить, исходя из своей теории. Конечно, не исключены и другие точки зрения.
Список литературы
Для подготовки данной работы были использованы материалы с сайта http://bio.1september.ru
Похожие работы
... когда какой цветок раскрывается, и прилетают именно к этому времени, вероятно, улавливая изменения в состоянии физических полей в окружающей их природе. Так же как и в неживой природе, электрическое и магнитные поля живого организма взаимосвязаны. В крови животных и человека обнаружен биогенный магнетит, который, по-видимому, позволяет живому организму чувствовать изменения магнитного поля Земли ...

... процессах. Главной задачей органической химии является анализ и синтез веществ, образующихся в биологических системах, живых организмах. Отсюда вытекает тесная связь химии и физики с другим разделом естествознания, с биологией. Изучение живых организмов позволяет увидеть множество чисто физических явлений: циркуляцию и гидродинамику протекания крови, давление в сосудах и т.д. Биология - очень ...
... и механицистской – мы сталкиваемся сразу с несколькими парадоксами. «Механицисты» рассматривают то человекоподобную машину, то машиноподобного человека; «органицисты» же видят в человеке то «воплощение чаяний» природы, то, всего-навсего, одно из природных созданий, не обладающее особыми привилегиями по сравнению со всеми остальными. Хорошо известно, что в эволюции живых организмов на Земле ...
... эти формы служат лишь для поддержания постоянного состава внутренней среды (гомеостаза) и для защиты открытых участков тела, непосредственно граничащих с внешней средой (например, жабр у пецилии [9]). Голые твари, одетые в незримую “одежду” из АФК, могут приносить пользу не только самим себе, но и всему водному биоценозу в целом: генерируемые ими активные формы кислорода освежают воду, ...
0 комментариев