Навигация
3.3 Транспорт липидов
Перенос мембранных липидов от места их синтеза к месту назначения осуществляется при помощи двух процессов: 1) трансмембранного флип-флоп-перехода; 2) внутримембранного транспорта. Данные о скорости флип-флопа обсуждались в в связи с асимметрией мембраны. Скорость флип-флоп-перехода фосфолипидов особенно велика для тех мембран, в которых происходит биосинтез липидов; ее характерное время составляет величину порядка нескольких минут. Имеются данные о том, что этот процесс осуществляется при участии белков и, возможно, требует гидролиза АТР. Было также показано, что холестерол способен к быстрому спонтанному флип-флоп-переходу. Следовательно, транспорт через мембрану эндоплазматического ретикулума из цитозоля в просвет происходит довольно быстро.
В транспорте липидов от одной клеточной мембраны к другой участвуют несколько процессов. В разных случаях наиболее важным может оказаться какой-то один из них.
1. Самопроизвольный перенос липидов путем диффузии мономерных липидных единиц через водную фазу.
2. Диффузия липидов через постоянные или временные места соединения двух контактирующих мембран.
3. Транспорт с участием белков, катализируемый или белками, облегчающими высвобождение липидов из донорной мембраны, или липидсвязывающими белками.
4. Транспорт с участием везикул, при котором липиды, как и мембранные белки, транспортируются в ходе непрерывного отпочковывания и слияния с мембранами внутриклеточных везикул. Этот процесс может быть энергозависимым.
Рассмотрим вначале, что известно о самопроизвольной диффузии мембранных липидов между мембранами. Как показывают многочисленные исследования, липиды могут самопроизвольно перемещаться между моноламелляриыми везикулами или между фосфолипидными везикулами и биомембранамн. В большинстве случаев при этом происходит десорбция мономериых липидов с поверхности донорной мембраны и свободная диффузия через водную среду к акцепторной мембране. Лимитирующим этапом (по крайней мере при избытке акцепторных мембран) является высвобождение липидов из донорной мембраны. В этих условиях характерное время переноса зависит от величины свободной энергии десорбции. Ясно, что менее водорастворимые липиды (т. е. липиды с низкой критической концентрацией мицеллообразования) должны преодолевать при десорбции более высокий энергетический барьер, а следовательно, их перенос должен осуществляться медленнее. Скорость переноса зависит не только от гидрофобности переносимого липнда, но и от состава и физического состояния донорного бислоя.
Например, ганглиозид GMb находясь в фосфатидилхолиновых везикулах, существует в монодисперсном состоянии. Благодаря наличию гидрофильных полярных групп он не совершает флип-флоп-переходов через мембрану везикул, но характерное время его переноса везикулами составляет около 40 ч при 45 °С [153]. Напротив, нейтральные ганглиозиды, лишенные остатков сиаловой кислоты (например, асиало-GMi), образуют в везикулах гелеобразный кластер, и характерное время их переноса составляет около 500 ч. Смесь холестерола и фосфолипидов в везикулах тоже образует сложные фазы, и это может влиять на кинетику переноса холестерола. Стабилизация холестерола в мембране могла бы происходить за счет благоприятных взаимодействий со специфическими фосфо-липидами, например со сфингомиелином.
Перенос новосинтезированного холестерола из эндоплазматиче-ского ретикулума в плазматическую мембрану осуществляется всего за 10 мин. На процесс оказывают влияние агенты, блокирующие биоэнергетические реакции в клетке, например цианид. Эти и другие данные свидетельствуют о том, что внутриклеточный транспорт холестерола является энерогозависимым процессом и протекает при участии везикул. В принципе он может перевесить любой спонтанный перенос. Однако единого мнения на этот счет не выработано. Серьезной проблемой является то, что оценки доли холестерола, присутствующего в плазматической мембране, от общего его количества в клетке сильно варьируют (от 25 до 95%). На первый взгляд кажется, что количество холестерола, поступающего в некоторые клетки и выходящего из них, можно оценить, используя данные о скорости самопроизвольной диффузии мономеров, однако неясно, пригодны ли в данном случае механизм спонтанного переноса и указанные скорости.
Характерное время переноса фосфолипидов из фосфолипидных везикул гораздо больше, чем холестерола. Например, для дипальмитоилфосфатидилхолина оно составляет 83 ч при 37° в случае везикул из димиристоилфосфатидилхолина. Скорость переноса фосфолипидов с помощью этого механизма слишком мала, чтобы соответствовать реальным скоростям межмембранного транспорта.
3.3.1 Транспорт липидов у прокариот
Остановимся вначале на переносе фосфолипидов в случае грамотрицательных бактерий. Это относительно простая система, поскольку в ней имеются только две мембраны. Фосфолипиды синтезируются в плазматической мембране и должны транспортироваться в наружную мембрану. Имеются данные о том, что между этими мембранами осуществляется быстрый обмен липидами. Так, фосфатидилэтаноламин достигает наружной мембраны за характерное время - 3 мин. В этом процессе не участвуют белки, липиды или АТР, и каким-то образом зависит от протондвижущей силы. Липиды, внедрившиеся в наружную мембрану, могут также быстро перемещаться к внутренней мембране; это относится даже к фосфатидилхолину, который в норме не обнаруживается в Е. coli. Таким образом, процесс, по-видимому, не является специфическим. Тем не менее у мутантиого штамма с дефектной диацилглнцеролкиназой диацилглицерол накапливается в плазматической мембране и к наружной мембране не транспортируется.
Механизм липидного обмена между этими двумя мембранами неизвестен, но обычно полагают, что внутренняя и наружная мембраны имеют места соединения или области адгезии. По-видимому, именно через эти участки и происходит диффузия липидов. Здесь же могут концентрироваться белки, экспортируемые к наружной мембране.
Похожие работы
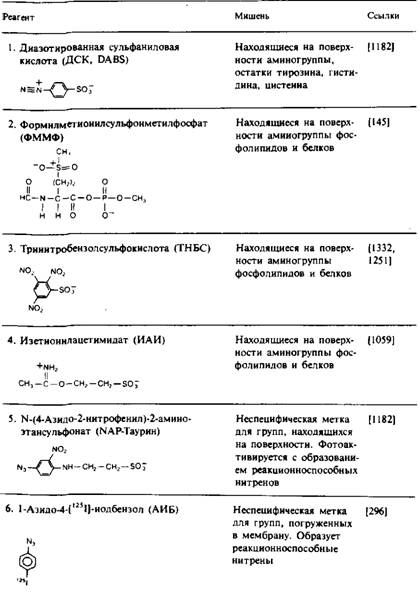



... и индуцируют трансмембранную миграцию липидных молекул, поэтому полученные результаты бывает трудно интерпретировать. Детальная оценка достоинств и недостатков методов изучения липидной асимметрии в мембранах дается, например, в обзорах. МЕТОДЫ УСТАНОВЛЕНИЯ ТРАНСМЕМБРАННОГО РАСПРЕДЕЛЕНИЯ ЛИПИДОВ Химическая модификация фосфолипидов Относительно легко подвергаются химической модификации только ...
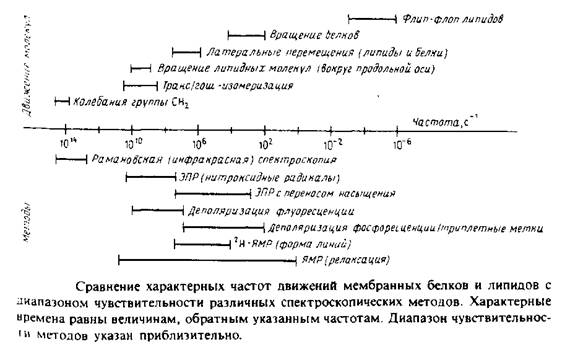
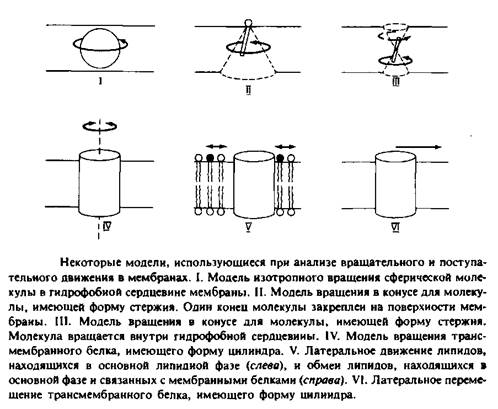
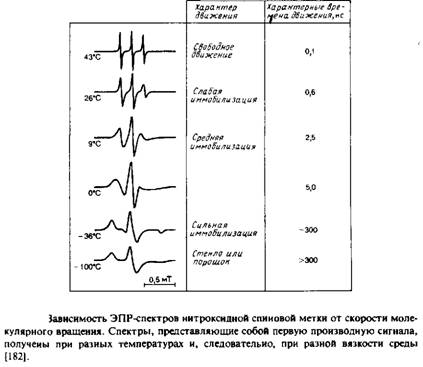
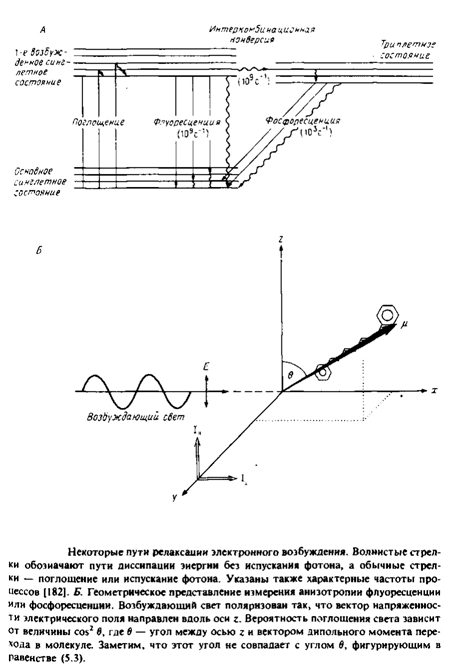
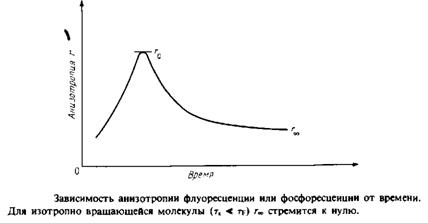
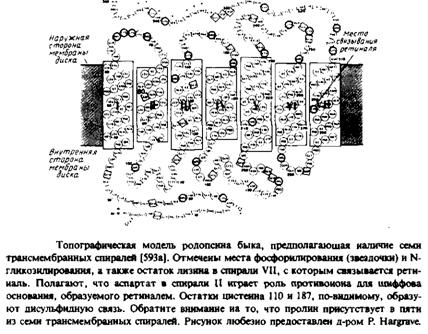
... к уменьшению латеральной диффузии. 5. Липидно-белковые взаимодействия Большинство методов, применяемых для изучения упорядоченности и динамических свойств мембран, используется и для исследования липидно-белковых взаимодействий. Работы по изучению этих взаимодействий были в основном направлены на выяснение влияния мембранных белков на физическое состояние липидов. Рассмотрим типичную мембрану ...
... является курсом, для изучения которого необходимо наличие знаний об основных принципах организации биологических молекул, строении и механизмах действия ферментов. Дисциплина биохимия мембран относится к дисциплинам специализации федерального компонента. 5. Распределение времени, отведенного на изучение дисциплины по учебному плану Форма учебной работы Форма обучения Очная По ...
... линейной последовательности нуклеотидов) явилось качественным переходом химической эволюции на свою следующую ступень или отправной точкой для самой ранней стадии биогенеза, непосредственно предшествующей процессу возникновения жизни. Следовательно, можно с определённой долей уверенности предположить, что стадия накопления „правильного“ для биогенеза химического потенциала сумела подготовить ...
0 комментариев