Навигация
Реферат
Белки нервной системы
ВВЕДЕНИЕ
Значительная часть белков нервной системы идентична белкам других органов и тканей в силу общности ряда базовых процессов жизнедеятельности. Однако существует обширная категория нейроспецифических белков, связанных с особым устройством и функциями нервной системы. Поскольку эта система функционирует как единое целое, невозможно в ряде случаев рассматривать только нейроспецифические белки, отвлекаясь от других белков. Можно лишь, стремясь акцентировать биохимические особенности нервной системы, уделить особое внимание нейроспецифическим белкам, не исключая из описания и некоторые другие белки в той мере, в которой это необходимо для полной характеристики белковых комплексов.
Специфичность белков для нервной ткани определяется критериями: а) наличием их преимущественно в нервной ткани, причем их количество должно существенно превышать таковое в остальных тканях животного организма, – условный, но общепринятый критерий; б) участием этих белков в реализации специфических функций нервной системы, например процессах генерации и проведения нервного импульса, установлении межклеточных контактов в нервной ткани, регуляции проницаемости ионных каналов, в механизмах обучения и формировании памяти; в) тесной взаимосвязью между биоактивностью нейроспецифических белков и функциональным состоянием нервной системы.
Изучение физико-химических свойств, локализации в отделах мозга, клетках и субклеточных структурах нервной ткани, особенностей метаболизма нейроспецифических белков или сроков появления их в процессе онтогенеза позволяет приблизиться к пониманию фундаментальных механизмов функционирования мозга. Установлена связь нейроспецифических белков с некоторыми патологическими состояниями организма, главным образом с развитием нервно-психических заболеваний. Обнаружение некоторых нейроспецифических белков в спинномозговой жидкости или сыворотке крови может рассматриваться в качестве индикатора повреждения нервной ткани.
Идентификация нейроспецифических белков может быть осуществлена различными способами:
1) сравнением белкового спектра мозга с белковыми спектрами других органов, в том числе путем наложения электрофореграмм после двумерного электрофореза; при этом могут быть выявлены как новые белки, характерные только для нервной ткани, так и их изоэлектрические точки, молекулярные массы, субъединичный состав и даже примерное количество;
2) с использованием иммунохимических методов, позволяющих определить нейроспецифические антигенные детерминанты, в том числе методом моноклональных антител и с помощью истощенных антисывороток; обработанные таким образом антисыворотки содержат антитела только к нейроспецифическим антигенным детерминантам;
3) с помощью направленного поиска нейроспецифических белков в различных участках и отделах мозга, в клеточных популяциях и в субклеточных структурах;
4) с помощью направленного поиска нейроспецифических изоферментов путем выявления ферментативной активности уже известных ферментов у вновь выделенных нейроспецифических белков;
5) с использованием методов генной инженерии, когда в качестве исходного материала применяется м-РНК мозга, с которой транскрибируется характерный нейроспецифический белок;
6) посредствам «дедуктивного» определения аминокислотных последовательностей белков нервной ткани – по нуклеотидным последовательностям генетической ДНК и м-РНК.
К настоящему времени различными методами идентифицировано более двух сотен нейроспецифических белков, однако информация о большинстве из них сводится в основном к сообщению об их выявлении и описанию ряда физико-химических и антигенных свойств. Представлены примеры наиболее изученных их них, классифицированных по функциональным и химическим характеристикам. В особых случаях, когда это полезно для восприятия путей познания биохимии мозга, приведены сведения об истории их открытия и изучения. В частности, описание белков, модулирующих состояние мембран и эффекты ионов Са+, неслучайно представлено первым, так как к ним относится первый из открытых и обстоятельно изученных нейроспецифических белков – S‑100.
1. НЕФЕРМЕНТНЫЕ НЕЙРОСПЕЦИФИЧЕСКИЕ СА+-СВЯЗЫВАЮЩИЕ БЕЛКИ
Очень многие белки ЦНС так или иначе взаимодействуют с ионами Са. Однако особо выделяют группу белков с очень высоким сродством к Са+, которые регулируют перемещения и концентрации Са+ и, благодаря способности менять конфор-мацию при связывании Са+, участвуют в разнообразных специфических процессах. Многие из белков этой группы называют калбиндинами. По особенностям структуры различают ан-нексины, содержащие длинные консервативные последовательности аминокислот, преимущественно дикарбоновых, и белки, обладающих так называемой «EF-pyKoff – петлей из 12–14‑и аминокислот, образующих как бы гнездо для Са+, фланкированные а-цепями.
К аннексинам относится первый открытый нейроспецифический белок – S‑100. Белок S‑100, точнее, как было установлено позже, – группа белков S‑100, был открыт в 1965 г. Б. Муром и Мак-Грегором при сравнении белковых карт водорастворимых белков мозга и печени. После хроматографии и электрофореза был выявлен первый специфический белок нервной ткани, названный белком Мура или белком S‑100, поскольку он остается в растворе при 100%-ном насыщении Н2S04 при рН 7,2. В дальнейшем белокБ‑100 был выделен в препаративных количествах из головного мозга человека, обезьяны, собаки, кролика, свиньи, крысы, мыши. В других тканях животных этих же видов – печени, почках, мышцах, в эритроцитах и сыворотке крови – он практически отсутствовал. Установлено, что S‑100 может содержаться в других органах и тканях, но в количествах, в 10-10 раз меньших, чем в нервной ткани. Интересно, что в 1984 г. белок S‑100 был обнаружен японскими исследователями в жировой ткани, из которой он высвобождался при действии адреналина in vitro. Кроме того, его присутствие иммунологически выявлено на поверхности клеток Лангерганса и родственных им клеток лимфоузлов.
S‑100 является гетерогенным кислым Са-связывающим белком. Он состоит из двух главных фракций: S‑100 А и S‑100 В, субъединичный состав которых соответственно аа и ар. Интересно, что аминокислотная последовательность р-субъединицы близка к таковой других Са-связывающих белков.
В зависимости от способа выделения может быть выявлено различное количество фракций и подфракций этого белка, что отражает как его природную гетерогенность, так и различные артефакты, связанные с методами выделения. Например, при электрофорезе с использованием высокой концентрации ПААГ обнаруживалось 5 фракций, причем все они реагировали с антисывороткой к белку S‑100. На сефадексе G‑100 белок S‑100 может быть разделен также на 5 фракций, обозначаемых как f1? f2, f3, f4, f5> причем до 85% этого белка приходится на долю первой фракции. Последующие стадии очистки приводят к разделению этой первой фракции на подфракций f1A, f]B, fjri основная масса белка была сосредоточена в последней подфракций, молекулярная масса которой составляла 19–22 кД. Кроме указанных субфракций в эту же группу включают в настоящее время еще около десятка родственных белков, содержание которых относительно невелико.
Своеобразен аминокислотный состав белка S‑100: характерно высокое содержание кислых аминокислот – около 36% приходится на остатки глутаминовой и 22% – на остатки аспарагиновой кислоты, т.е. более половины аминокислотного состава белка приходится на моноаминодикарбоновые аминокислоты. Этим определяются кислые свойства и низкая изоэлектрическая точка белка S‑100.
Из оставшихся 42% аминокислотных остатков основная масса приходится на гидрофобные алифатические аминокислоты, которые придают глобулам белка S‑100 частично гидрофобный характер. Наконец, можно отметить, что 3–4% от общего аминокислотного состава приходится на цистеин. Часть SH-rpynn цистеина свободна и способна к взаимодействию с ионами Са+. Такое взаимодействие приводит к значительному изменению конформации молекул белка S‑100. Меняется пространственное расположение гидрофильных и гидрофобных участков. В конечном счете изменяется способность S‑100 к миграции через мембраны клетки.
Белок S‑100 сосредоточен преимущественно в астроцитах – до 85–90% от общего содержания в нервной ткани. В олигодендроцитах его количество невелико. В нейронах обнаружено не более 10–15% от общего количества белка S‑100.
С помощью радиохимических, цитохимических и иммунохимических методов установлена внутриклеточная локализация белка S‑100. Основная масса этого белка сосредоточена в цитоплазме клеток и 15% – в мембранных структурах: в пре- и постсинаптических мембранах, ядерной мембране и плазматической мембране олигодендроглии. В ядрах нейронов его содержание крайне мало, несколько больше белка S‑100 найдено в ядрышках.
В то же время, недавно обнаружено, что в поясничном отделе спинного мозга крыс а-субъединица белка S‑100 локализована преимущественно в нейронах, а р-субъединица – в сателлитных глиальных и шванновских клетках.
Интересен вопрос об интенсивности биосинтеза белка S‑100 и появлении его в структурах мозга в онтогенезе. В мозге эмбриона человека он появляется на 10–15 неделе в мозжечке, Варолиевом мосту, стволе мозга, среднем и спинном мозге. К концу 30‑й недели происходит отчетливое накопление белка S‑100 во всех отделах ЦНС, кроме лобной доли коры больших полушарий, где повышение количества этого белка совпадает во времени с появлением биоэлектрической активности мозга.
Подробно изучено накопление белка S‑100 на различных этапах онтогенеза у грызунов. Показано, что в мозге мышей с 3‑го до 15‑го дня постнатального развития уровень этого белка остается относительно низким, а с 16‑го до 22‑го дня происходит быстрое возрастание его содержания примерно в 4 раза.
Содержание белка S‑100 в мозге повышается при обучении, тренировках и формировании условных рефлексов у животных. В период обучения происходит усиление биосинтеза белка S‑100, что подтверждается более интенсивным включением в него меченых аминокислот. Известный нейрохимик Х. Хиден и его сотрудники обнаружили, что наиболее интенсивный биосинтез данного белка происходит в пирамидальных клетках гиппокам-па. При интрацистернальном введении антисыворотки к белку S‑100 процесс обучения у животных нарушается.
Корреляция количества белка S‑100 в головном мозге со способностью к обучению показана и иным способом. У мышей некоторых инбредных линий, характеризующихся лучшей обучаемостью, содержание S‑100 выше.
Однако вопрос о непосредственном участии белка S‑100 в формировании и хранении памяти нельзя считать окончательно решенным. Не исключена возможность, что его участие является опосредованным.
На основании экспериментального материала и косвенных данных выдвинуто несколько предположений о возможных молекулярных механизмах участия белка S‑100 в специфических функциях нервной системы. Большинство авторов отдает предпочтение гипотезе о роли упомянутых выше конформационных изменений молекул белка S‑100, наступающих при взаимодействии его SH‑групп с ионами Са+ с последующим возрастанием на поверхности белковой глобулы количества гидрофобных групп. При проведении нервного импульса важным лимитирующим фактором служит проницаемость ионных каналов; в присутствии свободных ионов Са+ ряд каналов становится непроницаемым для ионов К+ и Na+. В этом случае функциональная роль белка S‑100, по-видимому, связана с регуляцией проницаемости ионных каналов посредством связывания свободных ионов Са+.
В нервной ткани велико содержание кальмодулина – одного из важнейших регуляторов и посредников эффектов Са+. Он присутствует и в других тканях и включение его в категорию нейроспецифических белков условно, однако его роль в нервной ткани велика: он участвует в активации Са+-ионами многих ключевых протеинкиназ и ряда других ферментов. Относительно низкомолекулярный белок – 17 кД – он консервативен по первичной структуре, высокостабилен и содержит четыре центра связывания Са*. Интересно, что активность кальмодулина подавляется хлорпромазином – одним из нейролептиков, применяемых при подавлении синдрома шизофрении.
В свою очередь функции кальмодулина контролируются, по крайней мере, двумя белками. Первый – кальцинейрнн. Он состоит из двух субъединиц – 15 и 61 кД и, обладая высоким сродством к кальмодулину, ингибирует его активность. Кроме того, кальцинейрин обладает протеинфосфатазной активностью и как бы обращает результаты действия протеинкиназ, включаемых кальмодулином.
Второй белок, способный связывать и как бы резервировать кальмодулин, является липопротеином. Это так называемый фосфомиристин В его молекулу входит жирная кислота – миристиновая. Кроме того, в нем велика доля гидрофобных аминокислотных остатков. Все это определяет его способность встраиваться в мембраны. В то же время он может фосфорилироваться под действием протеинки-назы С. В дефосфорилированном состоянии он связывает, резервирует кальмодулин, а после фосфорилирования – освобождает его. Содержание его в ткани мозга также велико.
Неизвестны пока функции другого связывающего кальций белка сиалогликопротеина GP‑350. Он имеет небольшую молекулярную массу – 11,6 кД; характерной его особенностью является высокое содержание остатков глутаминавой и аспарагиновок кислот, что и обусловливает взаимодействие с ионами Са». Растворимая форма гликопротеина GP‑350 сосредоточена в перикарионе нейронов и в аксонах; иммунологическая идентичная мембранная форма обнаружена в синаптосомах.
Обширные исследования посвящены функциям и свойствам мембранного нейроспецифического белка В‑50. В‑50 – один из основных фосфорилируемых белков плазматических мембран синапсов. Иммунохимически показано, что он локализован преимущественно в пресинаптических мембранах. Молекулярная масса белка 48 кД. Он является эндогенным субстратом диа-цилглицерол-зависимой и Са+-зависимой протеинкиназы С. Активаторы протеинкиназы С стимулируют процесс синапти-ческой передачи в срезах гиппокампа. Фосфорилирование белка В‑50 приводит к относительно продолжительному изменению заряда и состояния каналов постсинаптической мембраны и состоянию «проторенности» синапса. Одной из причин этого может быть влияние фосфорилированного В‑50 на метаболизм фосфоинозитидов и, таким образом, на отношение белок/ли-пид в синаптической мембране. Интересно, что в процессе старения интенсивность такого фосфорилирования снижается, чем, возможно, обусловлено снижение пластичности синапсов при старении.
Особо следует отметить высокую чувствительность процесса фосфорилирования белка В‑50 к адренокортикотропину, неодинаково выраженную в разных структурах мозга. Так, АКТГ1-24 в 10 раз более эффективен в торможении фосфорилирования белка В‑50 в синаптических мембранах из септальной области мозга, чем в мембранах из целого мозга. На основании этих наблюдений сделано заключение об участии белка В‑50 в функционировании пептидергических синапсов.
В процессах: синаптической передачи принимает участие еще один нейроспецифический белок – фодрин. Это – структурный белок постсинаптических мембран глутаматергических синапсов. Молекулярная масса его очень велика – 230 кД. Функциональная роль фодрина связана с тем, что он блокирует рецепторы глутамата. Г. Линч и М. Бодри предложили гипотезу, согласно которой повышение концентрации ионов Са+ вблизи постсинаптической мембраны активирует мембранную Сй-зависимую протеиназу калпеин, которая расщепляет фодрин. Результатом этого является освобождение активных рецепторов глутамата, ранее экранированных фодрином, и повышение проводимости синапса, которое наблюдается в течение 3–6 суток.
Недавно методом иммунохимического скрининга с помощью моноклональных антител к компонентам поверхности синаптосом идентифицирован новый фосфопротеин F 1–20, который локализуется в синапсах головного мозга, причем его содержание начинает резко возрастать после 7 дня постнатальной жизни животного. Показано, что этот фосфопротеин является так же, как и фодрин, субстратом для калпеина нейронов.
Некоторые нейроспецифические белки, модулирующие состояние мембран, не являются кислыми белками. Часть их обладает выраженными катионными свойствами. В течение последнего десятилетия были основательно изучены так называемые синапсины. Они составляют 0,2–0,4% от общего белка мозга и образуют целое семейство фосфопротеинов – la, lb, Па, ИЬ – с молекулярным весом 55000–86000 и изоэлектрической точкой в зоне рН 10,5 и 6,7–6,9. Процессы фосфорилирования-дефосфорилирования синапсинов тесно сопряжены с функциями везикул в нервном окончании. Де-фосфорилированный синапсин связывается с мембранами везикул и повышает их сродство к актиновым филаментам. Везикулы, вступившие в соединение с актином, образуют недеятельный, резервный пул. Фосфорилирование синапсинов, происходящее при повышении концентрации Са+ в терминали с помощью кальмодулин-зависимой протеинкиназы, снижая сродство синапсинов к мембранам везикул, ведет и к уходу их от актиновых филаментов и облегчает «плавление» везикул, необходимое для выброса медиатора.
После того как в результате фосфорилирования синапсина везикула переходит из «резерва» в активное состояние, целый каскад нейроспецифических белков обеспечивает ее контакт с пресинаптической мембраной, плавление и выход медиатора. Среди них опять-таки ключевая позиция принадлежит Са*+; он воздействует на сложный комплекс синаптических белков – синаптобревин, синалтофизин, синтаксин, синаптогамин, на фос-фолипиды мембран, регулирующих плавление везикулы, и, наконец, на синаптопорин, который, собственно, и формирует пору, канал, через который истекает содержимое везикулы. Детальные характеристики этих белков еще требуют уточнения. Интересно, однако, что именно они повреждаются под действием самых мощных из известных токсинов – столбнячного и ботулинического.
Похожие работы

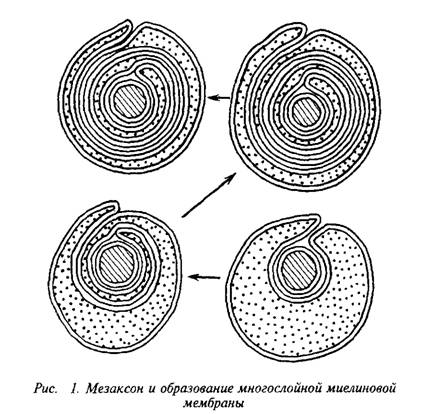
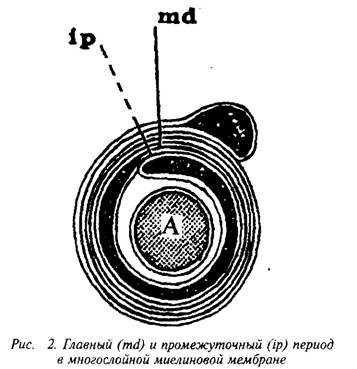
... и инозитолтрифосфат подвергаются химическим превращениям, требующим АТФ и ЦТФ и приводящим к восстановлению три-фосфоинозитида. Таким образом, цикл замыкается и уровень полифосфоинозитидов в мембране восстанавливается. 7. МИЕЛИН В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ Мозг человека содержит 120 г миелина, что составляет одну треть его сухой массы. Миелин – уникальное образование, организация которого ...
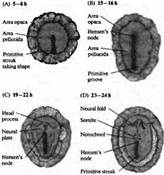



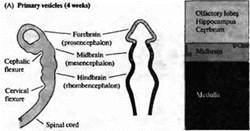
... уровне направленные мутации, что открывает дорогу к обнаружению важных генов позвоночных, которые могут иметь гомологов у некоторых видов более просто устроенных беспозвоночных. Развитие нервной системы в раннем периоде В раннем периоде морфогенеза позвоночных область гаструлы, из которой в дальнейшем формируется нервная система, представляет собой простую полоску эктодермы (рис. 1). ...
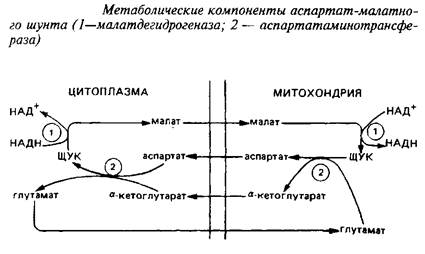
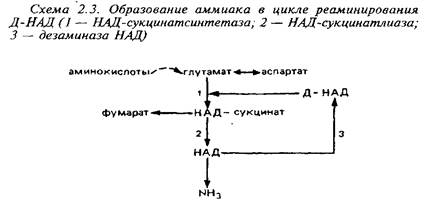
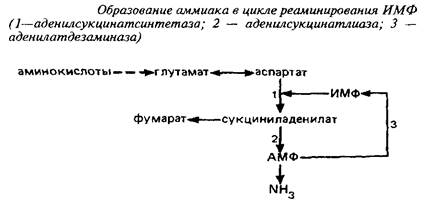
... видов животных. Региональные различия в концентрации метионина невелики. Влияние диеты на концентрацию метионина в мозге также незначительно из-за конкурентных отношений с нейтральными аминокислотами за транспортные системы. Метионин в пуле свободных аминокислот утилизируется на 80% для синтеза белка. Метаболизм свободного метионина до цистеина начинается с образования S-аденозилметионина, ...
... в синапсах вызывают деполяризацию или гиперполяризацию постсинаптической клетки. Потенциалы действия, представляющие собой короткие деполяризационные сигналы большой амплитуды, проводят по отросткам нейрона информацию из одного отдела нервной системы в другой. Все эти изменения мембранного потенциала вызваны движением ионов через клеточную мембрану. Например, направленное внутрь клетки движение ...
0 комментариев