Навигация
Синаптическая передача. Медиаторы
68465
знаков
0
таблиц
5
изображений
Синаптическая передача. Медиаторы
Введение
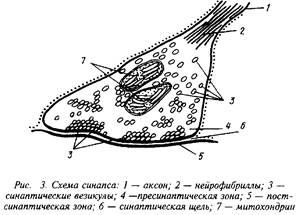
Ч. Шеррингтон обосновал представление об отсутствии межклеточной непрерывности в нервной системе и ввел понятие синапс для обозначения структуры, которая опосредует передачу сигнала от окончаний аксона к эффекторной клетке – нейрон), мышечному волокну, секреторной клетке железы. Синапс состоит из пресинаптического окончания и постсинаптической мембраны, разделенных синоптической щелью, которая заполнена рыхлым коллагеноподобным веществом.
Существуют два способа синаптической передачи – электрический и химический.
Возможно и сочетание обоих механизмов, электрического и химического, в одном смешанном синапсе, однако в нервной системе млекопитающих преобладают чисто химические синапсы.
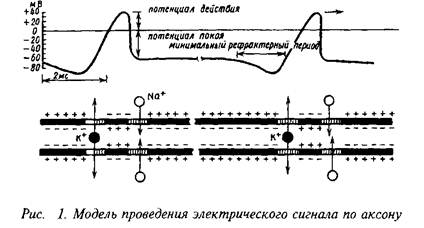
В электрических синапсах, количество которых в нервной системе относительно невелико, потенциал действия пресинаптических окончаний обеспечивает ток, который деполяризует постсинаптическую мембрану. Морфологическую основу электрической передачи составляет щелевой контакт, для которого характерны тесное прилегание пре- и постсинаптической мембран, большая площадь контакта этих мембран, наличие ультраструктур, снижающих электрическое сопротивление в области контакта, – своего рода каналов, организованных в ввде правильной сети между пре- и постсинаптической мембраной.
Электрофизиологическими критериями электрической синаптической передачи являются: 1) отсутствие синаптической задержки; 2) проведение возбуждения в обоих направлениях; 3) независимость от потенциала пресинаптической мембраны; 4) устойчивость к изменениям концентрации ионов кальция и магния в среде, к асфиксии, низкой температуре, некоторым фармакологическим воздействиям.
Функциональная роль электрических синапсов состоит в осуществлении срочной передачи сигналов, обеспечивающей синхронизацию электрической активности группы нейронов, например группы мотонейронов во время прыжковых движений лягушки или плавательных движений рыбы. Электрические синапсы обнаруживаются между нервными клетками, однотипными по структуре и функциям.
Эволюция нервной системы сопровождается уменьшением числа электрических синапсов в пользу другого способа передачи – химического. В химическом синапсе нервный импульс вызывает освобождение из пресинаптических окончаний химического посредника – нейромедиатора, который диффундирует через синоптическую щель и вступает во взаимодействие с белками-рецепторами постсинаптической мембраны, В результате происходит генерация постсинаптического потенциала.
Химический механизм синаптической передачи по сравнению с электрическим более эффективно обеспечивает основные функции синапса: 1) одностороннее проведение сигнала; 2) усиление сигнала; 3) конвергенцию многих сигналов на одной постсинаптической клетке, пластичность передачи сигналов.
Химические синапсы передают два вида сигналов – возбуждающий и тормозной. В возбуждающих синапсах нейромедиа-тор, освобождаемый из пресинаптических нервных окончаний, вызывает в постсинаптической мембране возбуждающий пост-синаптический потенциал – локальную деполяризацию, а в тормозных синапсах – тормозной постсинаптический потенциал, как правило, – гиперполяризацию. Снижение сопротивления мембраны, происходящее во время тормозного постсинаптического потенциала, ведет к короткому замыканию возбуждающего постсинаптического тока, тем самым ослабляя или блокируя передачу возбуждения.
В настоящей работе рассмотрены организация пресинаптического окончания, механизмы экзоцитозо. Ейромедиаторов, химическая природа и функциональные особенности медиаторов.
1. Морфофункциональная организация химического синапса. Квантовая теория освобождения нейромедиатора
Одним из самых детально изученных химических синапсов считается нервно-мышечное соединение скелетных мышц, в котором нейромедиатором служит ацетилхолин. Относительная доступность этого синапса и удобство для изучения позволили нобелевскому лауреату Б. Катцу в 50‑е годы провести мик-роэлектрофизиологические исследования, которые вместе с данными электронной микроскопии составили основу квантовой теории синаптической передачи, справедливой для самых разных химических синапсов.
Согласно этой теории процесс освобождения нейромедиатора складывается из отдельных элементарных реакций, каждая из которых представляет собой выход одного кванта нейромедиатора. Когда потенциал пресинаптической мембраны находится на уровне покоя, т.е. к пресинаптическим окончаниям не поступают нервные импульсы, кванты нейромедиатора тоже освобождаются, но спонтанно и с низкой скоростью. Ответом постсинаптической мембраны на отдельные кванты является возникновение миниатюрных постсинаптических потенциалов, в случае нервно-мышечного синапса они называются миниатюрными потенциалами концевой пластинки. Деполяризация пресинаптической мембраны во время нервного импульса ведет к практически синхронному освобождению большого количества квантов – до нескольких сотен. В результате возникает вызванный постсинаптический потенциалов нервно-мышечном синапсе он называется потенциалом концевой пластинки), который, в случае достижения пороговой амплитуды, ведет к генерации потенциала действия в постсинаптической клетке.
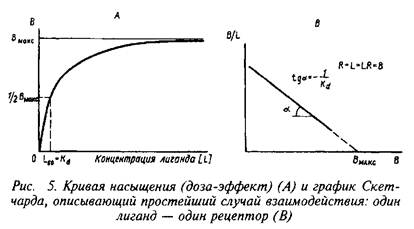
Освобождение индивидуальных квантов представляет собой статистический процесс: нервное окончание содержит п элементарных единиц, каждая из которых имеет независимую от других единиц вероятность ответа на нервный импульс. Если средняя вероятность ответа равна Ру то среднее число квантов, освобожденных в ответ на нервный импульс, определяется уравнением т=пР.
Повышение вероятности освобождения квантов при деполяризации пресинаптической мембраны связано с открыванием потенциал-зависимых Са+-каналов мембраны и входом Са в соответствии с их электрохимическим градиентом в пресинаптичское окончание. Таким образом, ионы Са участвуют в процессе электросекреторного сопряжения.
Квантовый характер синаптической передачи обусловлен тем, что нейромедиатор хранится в синаптических пузырьках. Присутствие везикул диаметром 40–200 нм, окруженных относительно плотной для электронов мембраной толщиной 4–5 нм, является характерной морфологической особенностью химических синапсов. Объем синаптического пузырька в двигательных нервных окончаниях диафрагмы крысы составляет 5,2–10нм. Такой пузырек может содержать порядка АЛ О молекул АХ, что вполне соответствует результатам оценки молекулярного состава кванта АХ – -10 молекул.
Синаптические пузырьки в аксоплазме нервного окончания сосредоточены в области, приближенной к пресинаптической мембране, около синаптической щели, причем пузырьки распределяются неравномерно, группируясь у периодически выступающих в аксоплазму утолщений пресинаптической мембраны – активных зон. По-видимому, в активных зонах находятся скопления потенциал-зависимых Са+-каналов, обеспечивающих вход Са+ в пресинаптическое окончание во время потенциала действия. В активных зонах обнаружены регулярно расположенные розеткообразные мембранные частицы диаметром около 15,0 нм, количество которых возрастает во время деполяризации пресинаптической мембраны. Можно предполагать, что эти частицы представляют собой точки слияния синаптических пузырьков с активной зоной, т.е. участки экзоци-тоза нейромедиатора в синаптическую щель.
При физиологических значениях частоты и длительности стимуляции нерва не удается выявить изменения количества синаптических пузырьков, несмотря на наблюдаемые изменения количества освобождаемого медиатора. Только при воздействиях, вызывающих истощение пресинаптических запасов медиатора, отмечается некоторый параллелизм изменений постсинаптических потенциалов и морфологических характеристик пресинаптических окончаний.
В период начального бурного освобождения медиатора, когда резко повышается частота спонтанных миниатюрных постсинаптических потенциалов, в области активных зон появляются многочисленные омегаподобные впячивания пресинаптической мембраны, которые соответствуют слипанию мембраны пузырька с участком освобождения. В фазу снижения квантового состава вызванных постсинаптических потенциалов наблюдается уменьшение количества синаптических пузырьков и увеличение площади поверхности пресинаптического окончания за счет встраивания мембран синаптических пузырьков в пресинаптическую мембрану. В дальнейшем идет процесс рециклизации синаптических пузырьков: от пресинаптической мембраны в нервное окончание отпочковываются мембранные структуры, которые сливаются, образуя окруженные мембраной цистерны; затем от цистерн отделяются синаптические пузырьки, вновь заполняемые медиатором, синтезированном в цитоплазме.
Процесс рециклизации синаптических пузырьков прослежен с помошью пероксидазы хрена. При добавлении этого маркера в среду инкубации электрическое раздражение нерва сопровождается усиленным захватом маркера синаптическими пузырьками и цистернами. Если после этого перенести препарат в среду, не содержащую пероксидазу хрена, и вновь подвергнуть его стимуляции, то пероксидаза освобождается в среду при экзоцитозе содержимого синаптических пузырьков.
Оценку динамики изменений запасов медиатора в пресинаптическом окончании во время ритмического раздражения нерва можно производить электрофизиологически путем измерения амплитуды и расчета квантового состава постсинаптических потенциалов. Классическими объектами для таких исследований стали нервно-мышечный синапс и симпатический ганглий. Структуры центральной нервной системы, где на теле или дендрите одного нейрона конвергируют много взаимодействующих синапсов, непригодны для таких оценок. Ритмическое раздражение двигательного нерва скелетной мышцы млекопитающего сопровождается быстрым снижением количества освобождаемого в ответ на каждый импульс медиатора, вплоть до некоторого относительно постоянного уровня. Это явление, называемое депрессией, показывает, что запас медиатора, способного к освобождению, ограничен и пополняется медленнее, чем расходуется. Депо АХ в нервно-мышечном синапсе оценивают примерной цифрой 210 квантов. Быстрая депрессия потенциалов концевой пластинки отражает расходование небольшой части депо АХ – так называемой фракции доступного медиатора, которая составляет примерно 10 квантов для нервно-мышечного синапса лягушки.
Интересной особенностью метаболизма пресинаптических окончаний является предпочтительная секреция вновь синтезированного в цитоплазме медиатора. После инкубации нервно-мышечного препарата или симпатического ганглия в среде с меченым предшественником АХ-холином ритмическое раздражение нерва приводило к секреции – АХ. Очевидно, существует фракция расположенных у активных зон синаптических пузырьков, которые после экзоцитоза их содержимого сразу же подвергаются рециклизации, заполняются только что синтезированным в цитоплазме медиатором и вновь готовы к экзоцитозу.
Следует отметить, что наряду с зависимой от ионов Са квантовой секрецией медиатора, которая обеспечивает передачу сигнала через синапс к постсинаптической клетке, а также происходит спонтанно, в отсутствие нервных импульсов, существует постоянная неквантовая утечка молекул медиатора из нервного окончания. В нервно-мышечном синапсе лягушки и млекопитающих неквантовая утечка создает концентрацию АХ в синаптической щели порядка 10~-10~М. Общее количество АХ, секретируемого неквантовым способом, превышает выход АХ, обусловленный спонтанной квантовой секрецией. Предполагается, что неквантовая секреция медиатора играет трофическую роль.
Нервно-мышечный синапс является экспериментальным объектом, удобным для исследования электрофизиологическими методами. Однако изоляция нервных окончаний и синаптических пузырьков для биохимического исследования крайне затруднена вследствие того, что двигательные нервные окончания составляют слишком малую долю от объема ткани скелетной мышцы. Гораздо более адекватным препаратом для изучения экзоцитоза нейромедиаторов являются синаптосомы – пресинаптические окончания, выделяемые из нервной ткани, как правило, вместе с веществом, заполняющим синаптическую щель, и участком постсинаптической мембраны. Деполяризация синаптосом физиологическим раствором с высокой концентрацией К+ вызывает зависимую от Са»^ секрецию нейромедиаторов.
Похожие работы

... В этом случае раздражения разделены интервалом времени, а не пространством и более высокую эффективность повторного раздражения можно считать результатом временной суммации. В современных учебниках по физиологии приводятся понятия временной и пространственной суммации, но теперь об их механизме известно больше, чем в то время, когда они были впервые описаны. Нервные волокна многократно ветвятся, ...

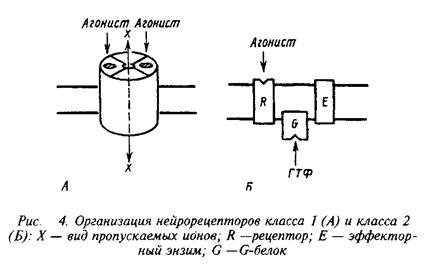
... роль нейрорецепторов сводится к созданию специфических информационных входов, организующих единый функциональный ансамбль нейронов. Именно совокупность рецепторов определяет лицо клетки и ее реакции на поступление разнообразных химических сигналов. Молекулярные механизмы, лежащие в основе модуляции эффективности синаптической передачи, в которых важную роль играют рецепторные процессы, имеют ...
... специфических сигналов. Представление о синапсах связано с Шеррингтоном (1935), высказавшим предположение о существовании специальных структурно-функциональных образований, обеспечивающих контакты между нейронами. Особенности рефлекторных реакций и некоторые свойства нервных центров обусловлены процессами, происходящими на синапсах. Синапс включает в себя три компонента: пресинаптический, ...
... в глутамат с образованием янтарного полуальдегида. HOOC-CH2-CH2-CH2-NH2 à HOOC-CH2-CH2-CH(O) ГАМК Янтарный полуальдегид ГАМК в организме γ–аминомасляная кислота является основным ингибиторным трансмиттером (передатчиком) в ЦНС. Она осуществляет нейрональную трансмиссию в 1/3 всех синапсов головного и спинного мозга. ГАМК играет важную роль в регуляции двигательных функций. При этом ...
0 комментариев