Навигация
Особенности пропионовокислого брожения: применение пропионовокислых бактерий
19832
знака
0
таблиц
1
изображение
Министерство сельского хозяйства РФ ФГОУ ВПО «Оренбургский государственный аграрный университет»
Реферат
по промышленной микробиологии
на тему
Особенности пропионовокислого брожения: применение пропионовокислых бактерий
Подготовил: студент
Акжигитов Абай Сарсенгалиевич
Оренбург - 2010
Содержание
Введение. Общая информация о пропионовокислых бактериях
1. Пропионовокислое брожение: химизм и особенности
2. Использование пропионовокислых бактерий в промышленности
Заключение
Список литературы
Введение: общая информация о пропионовокислых бактериях
Пропионовокислые бактерии - неспороносные грамположительные неподвижные палочки размером 0,5—0,8 или 1,0—1,5 мкм (в молодых культурах — искривленные, слегка ветвящиеся палочки, в более старых — кокковидной формы). Образуют колонии жёлтого, оранжевого или красного цвета, растут как в аэробных, так и в анаэробных условиях.
Пропионовокислые бактерии родственны по ряду свойств гетероферментативным молочнокислым бактериям Они, как и молочнокислые бактерии, не встречаются в почве или водоемах. Обитают в основном в рубце и кишечнике жвачных животных, в молочных продуктах (не в молоке). Пропионовокислые бактерии — возбудители пропионовокислого брожения, сбраживают глюкозу, лактозу и др. углеводы, а также некоторые спирты с образованием пропионовой и уксусной кислот и CO2. После молочнокислого брожения, когда лактоза превращена в молочную кислоту, начинают размножаться.
Пропионовокислые бактерии применяют для микробиологического синтеза витамина B12. Пропионовые бактерии могут синтезировать гемсодержащие белки. В их клетках обнаружены цитохромы.
Конструктивный метаболизм пропионовых бактерий претерпел дальнейшую эволюцию в сторону большей независимости от органических соединений внешней среды. Пропионовые бактерии характеризуются хорошо развитыми биосинтетическими способностями и могут расти на простой синтетической среде с аммонийным азотом в качестве единственного источника азота при добавлении к среде пантотеновой кислоты и биотина, а для некоторых видов и тиамина. У ряда пропионовых бактерий обнаружена способность к азотфиксации.
Местообитание пропионовых бактерий — кишечный тракт жвачных животных, молоко, твердые сыры, в приготовлении которых они принимают участие.
1. Пропионовокислое брожение: химизм и особенности
Основные продукты пропионовокислого брожения, вызываемого несколькими видами бактерий из рода Propionibacterium, — пропионовая (CH3CH2OH) и уксусная кислоты и CO2. Химизм пропионовокислого брожения сильно изменяется в зависимости от условий. Это, по-видимому, объясняется способностью пропионовых бактерий перестраивать обмен веществ, например, в зависимости от аэрации. При доступе кислорода они ведут окислительный процесс, а в его отсутствии расщепляют гексозы путём брожения. Пропионовые бактерии способны фиксировать CO2, при этом из пировиноградной кислоты и CO2 образуется щавелевоуксусная кислота, превращающаяся в янтарную кислоту, из которой декарбоксилированием образуется пропионовая кислота.
Основное энергетическое значение для пропионовокислых бактерий имеют так называемые ключевые реакции пропионовокислого брожения.
Под пропионовокислым брожением подразумевают биохимический процесс превращения бактериями сахара, молочную кислоту и ее солей в пропионовую кислоту. В этом брожении, кроме пропионовой кислоты, образуются и такие продукты, как уксусная кислота, углекислый газ, янтарная кислота, ацетоин, диацетил, другие летучие ароматические соединения - диметилсульфид, ацетальдегид, пропионовый альдегид, этанол и пропанол. Химизм данного брожения подобен типичному молочнокислому брожению с той разницей, что образовавшаяся молочная кислота в этом брожении не конечный продукт, а промежуточный. От других типов брожения пропионовокислое отличается высоким выходом АТФ, участием некоторых уникальных ферментов и реакций.
Пропионовокислым бактериям свойственен бродильный тип метаболизма: они расщепляют сахара по пути Эмбдена–Мейергоффа до пропионата, ацетата, СО2 и сукцината. Химизм пропионовокислого брожения хорошо изучен и описан.
В пропионовокислом брожении мы имеем дело с карбоксилированием пирувата, приводящим к возникновению нового акцептора водорода — ЩУК. Восстановление пировиноградной кислоты в пропионовую у пропионовокислых бактерий протекает следующим образом. Пировиноградная кислота карбоксилируется в реакции, катализируемой биотинзависимым ферментом, у которого биотин выполняет функцию переносчика CO2. Донором CO2-группы служит метилмалонил-КоА. В результате реакции транскарбоксилирования образуются ЩУК и пропионил-КоА:
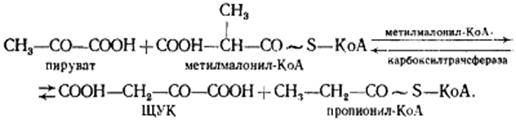
Ключевую реакцию брожения - превращение а-метилмалонил-КоА в сукцинил-КоА катализирует кофермент В12 (Ado Cbl).
Когда сбраживаемым субстратом является лактат, он сначала окисляется в пируват. Часть пирувата далее окисляется до ацетил-КоА и СО2, причем превращение ацетил-КоА в ацетат сопровождается образованием АТФ. Получение в процессе брожения окисленных продуктов, ацетата и СО2 уравновешивается сопутствующим восстановлением пирувата до пропионата.
Пировиноградная кислота - обязательное промежуточное соединение в брожении.
Пируват может быть превращен в пропионат несколькими путями:
1). пируват →акрилат→пропионат;
2). пируват→лактат→пропионат;
3). пируват + С 1 →сукцинат →метилмалонат →пропионат.
Первые две возможности у пропионовых бактерий не реализуются, и образование пропионата происходит из дикарбоновой кислоты по третьему пути.
Сначала лактат окисляется до пирувата при участии флавопротеида в качестве акцептора водорода, затем в реакции транскарбоксилирования образуется оксалоацетат. Донором СО2 служит (S)-метилмалонил-КоА, а переносчиком СО2 - биотин. Под действием малатдегидрогеназы и фумаразы образуется фумарат, который восстанавливается до сукцината в реакции, катализирующей фумаратредуктазой. Эта реакция сопряжена с синтезом АТР путем фосфорилирования, сопряженного с переносом электронов. Далее в КоА - трансферазной реакции образуется сукцинал-КоА. Затем под действием метилмалонил-КоА-мутазы, содержащей кофермент В12, осуществляется перегруппировка, ведущая к образованию (R)-метилмалонил-КоА, который, однако, не является субстратом для транскарбоксилазы. Скорее всего, (S)-стереоизомер образуется при действии специфической рацемазы. В зтом случае в реакции транскарбоксилирования синтезируется пропионил-КоА, и в результате последующего переноса КоА на сукцинат образуется пропионат.
В процессе сбраживания лактата в пропионат потребляется одна молекула NADH2. Она образуется при окислении лактата до ацетата в соответствии с суммарным уравнением реакций брожения:
Лактат + NADH2 + ADP + Pi → Пропионат + NAD + ATP.
Реакции, ведущие к образованию пропионовой кислоты у пропионовокислых бактерий, могут быть представлены следующей последовательностью:
1) Пируват + метилмалонил-КоА (а) ↔оксалоацетат+пропионил-КоА
2) Оксалоацетат+пропионил-КоА
3) Сукцинат + пропионил-КоА (+ КоА-трансфераза)↔сукцинил-КоА+пропионат
4) Сукцинил-КоА ↔(+метилмалонилизомеразая)↔метилмалонил-КоА
5) Метилмалонил-КоА (в) ↔(+метилмалонилрацемаза)↔метилмалонил-КоА(а)
Суммарно: пируват + 4H+→ пропионат.
Превращение оксалоацетата в сукцинат происходит в результате работы ферментов ЦТК: малатдегидрогеназы, фумаразы и сукцинатдегидрогеназы. Уксусная кислота образуется в результате окислительного декарбоксилирования пирувата:
6) Пируват + HAD+ + КоА(+пируватдегидрогеназа)↔ацетил-КоА+Н+ + НADH + CO2,
7) Ацетил-КоА + Фн (+фосфотрансацетилаза)↔ ацетил-Ф + КоА, ацетил-Ф + АDФ (+ацетилкиназа)↔ацетат + АТФ.
Итак, разобранный выше поток реакций приводит к синтезу пропионовой кислоты. Однако пропионовокислое брожение — более сложный процесс, поскольку наряду с пропионовой кислотой в качестве продуктов брожения образуются уксусная, янтарная кислоты и CO2 (см. рис. 1):
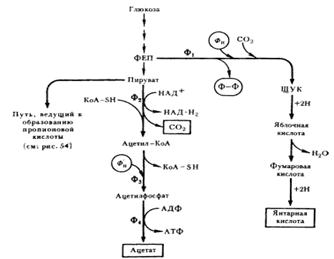
Рис. 1. Пути образования янтарной, уксусной кислот и CO2 пропионовыми бактериями: Ф1 — ФЕП-карбокситрансфосфорилаза; Ф2 — пируватдегидрогеназа; Ф3 — фосфотрансацетилаза; Ф4 — ацетаткиназа
Кроме основных продуктов в разных количествах в культуральной жидкости пропионовых бактерий обнаружены молочная, муравьиная, изовалериановая кислоты, этиловый и пропиловый спирты, уксусный и пропионовый альдегиды, ацетоин, диацетил. Состав конечных продуктов брожения зависит от культуры бактерий, состава среды и условий культивирования. Это касается как видов накапливаемых продуктов, так и количественных соотношений между ними.
Соотношение продуктов брожения может быть разное и в значительной степени зависит от степени окисленности источника углерода. При росте на среде с глицерином, например, отношение пропионовая: уксусная 2:1, с лактатом 1:1,5 и пируватом 1:2. С другой стороны, при культивировании пропионовокислых бактерий в строго анаэробных условиях соотношение между количествами кислот отклоняется в обратную сторону; именно в этом случае на одну молекулу уксусной всегда образуется 3 молекулы пропионовой. Также большое влияние имеет значение концентрации ионов водорода. При увеличении концентрации ионов водорода в среде изменяется соотношение основных продуктов брожения: образование уксусной кислоты увеличивается, а пропионовой заметно уменьшается. Соотношение пропионовой и уксусной кислот зависит от состава и свойств среды и внешних условий существования микроорганизмов. В сырах в период максимального развития культуры пропионовых бактерий (первая фаза) в основном образуются относительно окисленные соединения (уксусная кислота), в период спада развития – преимущественно более восстановленные (вторая фаза). Но при замедлении развития культуры P. shermanii (замена пептона аммонийными солями) уксусная кислота превалирует перед пропионовой, однако и в этом случае отношение пропионовой кислоты к уксусной возрастает. Этот пример служит доказательством связи продуцирования кислот с составом среды. Отношение пропионовой кислоты к уксусной зависит также от вида бактерий. В среде с глюкозой и дрожжевым автолизатом пропионовокислые бактерии рода P. thonii продуцируют указанные кислоты в отношении 5:1, для P. rubrum это отношение равно 3:1 и для P. shermanii – 2:1. Для культур P. shermanii (9 штаммов) отношения пропионовой кислоты к уксусной колеблется от 1,4 до 2,8.
Между количеством организмов и количеством образующихся кислот нет прямой связи. Изменение состава карбоновых кислот в питательной среде (соли молочной, пировиноградной и янтарной кислот) значительно влияет на продуцирование пропионовой и уксусной кислот культурами P. shermanii. Отношение пропионовой кислоты к уксусной изменяется, с лактатом оно равно 1,83 (17,55:9,55), с пируватом – 0,64 (8,20:12,75) и янтарной кислотой – 2,0 (3,30:1,50). В присутствии лактозы продуцирование пропионовой кислоты происходит более энергично, чем в присутствии глюкозы. При сбраживании культурами P.jensenii пировиноградной кислоты более окисленной, чем молочная, также наблюдается, что соотношение пропионовой и уксусной кислот смещается в сторону более окисленной уксусной. Соотношение кислот зависит от состава закваски. Так, при использовании закваски с двумя культурами – L. helveticum и Str. thermophilus это соотношение ниже, чем при использовании одной L.helveticum.
Таким образом, уникальность пропионовокислого брожения обусловлена: 1) участием ФЕП – карбоксилтрансфосфорилазы фермента, не обнаруженного у других организмов, синтезирующих пропионат; благодаря наличию этого фермента, брожение, осуществляемое пропионово-кислыми бактериями, работает как циклический процесс.
2) особым способом образования пропионата, которое сопряжено с восстановлением фумарата до сукцината и окислением пирувата до ацетата и СО2; транспорт электронов, сопровождающий эти реакции, сопряжен с окислительным фосфорилированием и синтезом АТФ;
3) высокий выход АТФ, превышающий выход АТФ в других известных брожениях. 1,5 М глюкозы могут дать пропионовокислым бактериям около 6 М АТФ.
Энергетическая эффективность пропионовокислого брожения связана также с выработкой пропионовыми бактериями новых метаболических способностей: реакций транскарбоксилирования и перегруппировки, участия в процессе КоА-производных. Образование дикарбоновой кислоты из пировиноградной с использованием механизма транскарбоксилирования вместо прямого карбоксилирования пирувата позволяет избежать дополнительных энергетических затрат на этом этапе брожения. Все это вместе взятое позволяет рассматривать пропионовокислое брожение как более совершенный из рассмотренных до сих пор способов получения энергии в анаэробных условиях.
Похожие работы
... картофель всплывает вследствие бурно идущего газообразования. По окончании брожения культуральную жидкость используют для исследования морфологии маслянокислых бактерий и качественного определения продуктов брожения. Качественная реакция на масляную кислоту. Получение маслянокислого железа (реакция с FeCl3). Нейтральные растворы маслянокислых солей при нагревании с FeCl3 приобретают коричневое ...
... продуктов, являются коагуляция казеина и гелеобразование (переход коллоидной системы молока из свободнодисперсного состояния, золя, в связаннодисперсное состояние - гель). Коагуляция казеина при производстве кисломолочных продуктов может осуществляться двумя способами - кислотным или сычужным. Кислотная коагуляция казеина вызывается молочной кислотой, которая накапливается в молочных продуктах ...
... процессы и протеолиз при созревании - в сыре увеличивается количество водорастворимых белков, пепгидов и аминокислот, а степень усушки сыра снижается. Классификация сыров и их технологические особенности По способу свертывания молока различают сыры сычужные и кисломолочные. Сычужные вырабатываются с помощью сычужного фермента, кисломолочные под действием молочной кислоты. Сыры сычужные ...
... - уксусную кислоту; некоторые гетероферментативные молочнокислые бактерии могут образовывать различные спирты, глицерин, маннит. Гетероферментативное молочнокислое брожение вызывают бактерии рода Lactobacterium и рода Streptococcus. Химизм этих брожений изучен не так хорошо, как спиртового или гомоферментативного молочнокислого брожения. Гетероферментативные бактерии образуют молочную кислоту ...
0 комментариев