Навигация
Основные закономерности передачи наследственных свойств
25578
знаков
0
таблиц
2
изображения
Реферат
«Основные закономерности передачи наследственных свойств»
1. Дигибридное и полигибридное скрещивание
Моногибридное скрещивание легко может быть получено в опыте. Однако в природных условиях скрещивание обычно происходит между особями, различающимися по многим признакам. Каковы же в этих более сложных случаях закономерности наследования? Чтобы ответить на этот вопрос, рассмотрим дигибридное скрещивание, т. е. наследование двух пар признаков. В качестве примера обратимся вновь к разным сортам гороха, изученным Менделем. Исходными формами для скрещивания взяты, с одной стороны, сорт гороха с желтыми и гладкими семенами, с другой— с зелеными и морщинистыми. В этом скрещивании мы имеем дело с двумя аллелями. Одна аллель включает гены окраски семян (желтая, зеленая), вторая — формы семян (гладкая, морщинистая). Если для скрещивания взяты гомозиготные формы, то все потомство в первом поколении гибридов будет обладать желтыми гладкими семенами. Следовательно, в первой аллели доминантной (как это уже известно, нам из анализа моногибридного скрещивания) окажется желтая окраска, рецессивной — зеленая (аллель А—а). Во второй аллели (обозначим ее B—b) гладкая форма семян доминирует над морщинистой. При самоопылении или скрещивании между собой гибридов первого поколения произойдет расщепление. По фенотипу получится четыре группы особей в различных численных отношениях: на девять желтых гладких (АВ) будет приходиться три желтые морщинистые (Ab), три зеленые гладкие (аВ) и одна зеленая морщинистая (ab). В кратком виде это расщепление можно представить следующей формулой: 9АВ:ЗAb:ЗаВ:1ab
Рассмотрим более подробно ход скрещивания и расщепления. Пользуясь принятыми символами, генотипы исходных гомозиготных родительских форм следует обозначить как ААВВ и aabb. Очевидно, исходя из гипотезы чистоты гамет, их половые клетки должны нести по одному гену от каждой аллели, т. е. гаметы будут у одной родительской формы АВ, а у второй — ab. В результате оплодотворения получится гибрид наследственного состава АаВb. Этот гибрид гетерозиготен по двум аллелям, но так как у него присутствуют гены А и В, то по фенотипу он сходен с одним из родителей. Результаты расщепления во втором поколении можно предсказать, если знать, какие гаметы получаются у дважды (по обеим аллелям) гетерозиготных гибридов первого поколения. Так как в гамете из каждой аллели может присутствовать только один ген (гипотеза чистоты гамет), то, очевидно, у двойных гетерозигот должны быть четыре сорта гамет, а именно: АВ, АЬ, аВ и ab. Встреча между любыми двумя из этих гамет, принадлежащих различным родителям, одинаково вероятна. Из четырех по два может быть 16 различных комбинаций. Все они представлены на таблице, где выписаны также все 16 образующихся при этом генотипов. Во всех 16 квадратах нарисованы также фенотипы соответствующих особей. Легко подсчитать окончательный результат расщепления F2, который уже приведен выше.
Когда скрещиваются организмы, различающиеся по признакам, в отношении которых наблюдается полное доминирование, то число возникающих во втором поколении гибридов различных генотипов значительно больше, чем число разных фенотипов. Как было показано, при дигибридном расщеплении наблюдается четыре разных фенотипа. Большинство их слагается из нескольких генотипов. Среди растений гороха, обладающих желтыми и гладкими семенами (фенотип АВ), как бы скрывается четыре разных генотипа, а именно; формы гомозиготные (ААВВ), гетерозиготы по признаку окраски семян (АаВВ), гетерозиготы по признаку формы семян (ААВb) и, наконец, формы гетерозиготные по обеим аллелям (АаВb). Таким образом, этот фенотип включает четыре разных генотипа. Растения с желтыми морщинистыми семенами (фенотип Аb) представлены двумя генотипами: гомозиготами ААbb и гетерозиготами Aabb. Два генотипа скрываются и за фенотипом с зелеными гладкими семенами (аВ), а именно ааВВ и ааВb. Что касается рецессивных форм с морщинистыми зелеными семенами, то они всегда гомозиготны и представлены одним генотипом aabb. Таким образом, число различных генотипических комбинаций у второго поколения гибридов (F2) оказывается равным девяти.
Рассмотренные количественные отношения между числом различных фенотипов и генотипов в F2 при дигибридном скрещивании справедливы для аллелей с полным доминированием. Ясно, что в случаях промежуточного характера наследования число фенотипически различных форм будет больше. Если по 6 обоим признакам наследование промежуточное, то количество фенотически различных групп равняется числу генотипически различных групп.
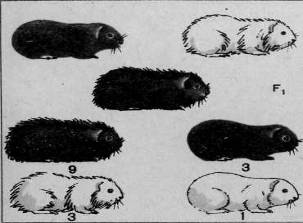
Рис. 1. Скрещивание и ход расщепления в двух поколениях двух пород морских свинок, различающихся по двум аллелям – окраске и форме шерсти (дигибридное скрещивание).
Ход дигибридного скрещивания можно показать и на примере животных. На рисунке 1 изображено дигибридное скрещивание двух пород морских свинок: черных гладких с белыми мохнатыми. В данном случае черная окраска доминирует над белой, мохнатая шерсть — над гладкой. Из рисунка без дальнейших пояснений ясен ход скрещивания, который совершенно аналогичен рассмотренному дигибридному скрещиванию горохов.
Второй закон Менделя. Сопоставим результаты дигибридного и моногибридного скрещиваний. Если учитывать результаты расщепления по каждой аллели в отдельности, то легко видеть, что соотношение, характерное для моногибридного скрещивания, сохраняется. При рассмотренном выше дигибридном расщеплении у горохов отношение числа желтых семян (А) к зеленым (а) равняется 12:4 (3:1). То же касается и отношения гладких семян (В) к морщинистым (b). Таким образом, дигибридное расщепление представляет собой, по существу, два независимо идущих моногибридных, которые как бы накладываются друг на друга. Это может быть выражено алгебраически как квадрат двучлена (3+1)2 = 32 + 2хЗ+12, или, что то же самое, 9+3 + 3+1. Мы подошли, таким образом, к формулировке второго очень важного закона, установленного Менделем, который можно назвать законом независимого распределения генов. Он гласит, что расщепление по каждой паре признаков (по каждой аллели) идет независимо от других пар признаков (относящихся к другим аллелям).
Цитологические основы дигибридного расщепления. Как связать закономерности дигибридного расщепления с теми процессами, которые совершаются в половых клетках при их созревании и оплодотворении? Эти отношения поясняются на схеме (рис. 106). Диплоидный набор хромосом представлен здесь двумя гомологичными парами. В парных хромосомах расположены аллельные гены. В палочковидных хромосомах гены А (красные) и а (синие), в сферических хромосомах гены В (красные) и b (синие). В результате мейоза из каждой гомологичной пары хромосом в гаметах остается по одной (см. схему). В результате оплодотворения в двойной гетерозиготе АаВb в каждой паре хромосом будут разные гены одной аллели (на схеме красная и синяя). При редукционном делении у гибрида первого поколения (F1) в равном количестве образуется четыре сорта гамет. Это зависит от того, что при мейозе во время конъюгации хромосом, предшествующей их расхождению, взаимное расположение хромосом разных гомологичных пар носит случайный характер. Если, например, к одному полюсу отходит «синяя» палочковидная хромосома, то из другой пары с одинаковой долей вероятности может отойти или тоже «синяя», или же «красная». В результате оплодотворения и развития второго поколения гибридов (F2) одинаково вероятно образование 16 категорий зигот. На схеме все возможные комбинации хромосом в зиготах врисованы в квадрате.
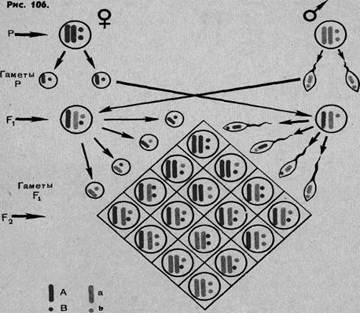
Рис. 2. Цитологические основы дигибридного скрещивания. Хромосомы, несущие доминантные гены, красные, рецессивные – синие.
Зная, что аллельные гены локализованы в гомологичных хромосомах, мы можем несколько иначе изобразить ход дигибридного скрещивания и расщепления, чем мы это делали до сих пор, представив в формулах гомологичные хромосомы в виде черточек.
Анализирующее скрещивание. Все изложенное выше о закономерностях в характере наследования признаков ясно показывает, что по фенотипу организма нельзя судить с достаточной полнотой о его наследственной структуре — его генотипе. Например, горох с желтыми гладкими семенами может быть гомозиготным (генотип ААВВ), а может быть и дигетерозиготным (АаВb) или гетерозиготным по одной аллели (ААВb и АаВВ). Определить генотип можно лишь по характеру расщепления гибридного поколения. Определение генотипа не только представляет теоретический интерес, но также имеет и большое практическое значение при селекционной работе по выведению или улучшению пород и сортов.
При этом широко используется анализирующее скрещивание, которое представляет собой скрещивание особи, генотип которой мы хотим определить, с формой чисто рецессивной по изучаемым аллелям. Такое скрещивание имеет большие преимущества, заключающиеся в том, что позволяют в первом же поколении гибридов определить гаметы анализируемой формы. Действительно, чисто рецессивная форма всегда гомозиготна. Например, зеленые морщинистые семена гороха имеют генотип ааbb и дают гаметы только одного вида — ab. Допустим, что горох, обладающий зелеными морщинистыми семенами, мы скрестили с горохом с желтыми и гладкими семенами, генотип которых нам неизвестен, и получили потомство, в котором 25% растений обладают желтыми гладкими семенами, 25% — желтыми морщинистыми, 25% — зелеными гладкими и 25% — зелеными морщинистыми. Очевидно, можно утверждать, что она образовывала 4 сорта гамет в равных количествах, т. е., другими словами, была гетерозиготной по двум аллелям.
Допустим, что в другом аналогичном скрещивании растений с теми же признаками мы не получили в потомстве никакого расщепления и все растения оказались имеющими лишь доминантные признаки по изучаемым аллелям (т. е. желтые гладкие семена). Это будет указывать на то, что взятая нами особь была доминантной гомозиготной (ААВВ). Этот случай уже был разобран подробно выше (54).
Похожие работы
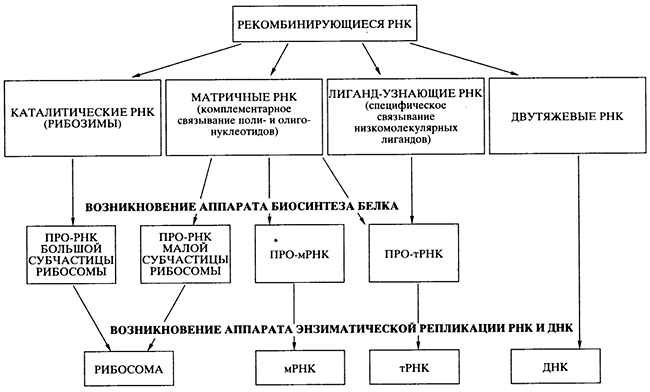
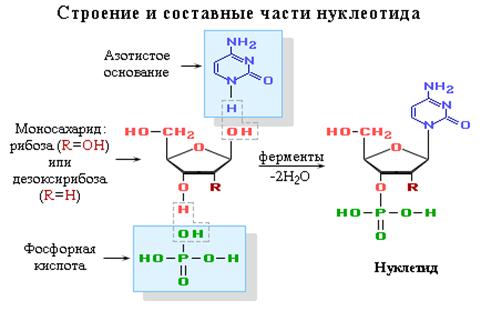
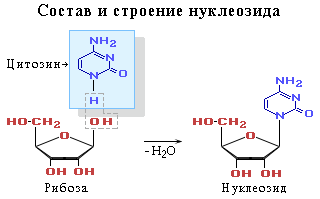
... , образуя вначале нуклеосому, а после гетерохроматин, из которого состоят хромосомы; ü передача наследственного материала происходит путем репликации ДНК; ü реализация наследственной информации в процессе синтеза белка. 5. РНК 5.1. Состав РНК Первые сведения о нуклеотидном составе РНК относились к препаратам, представляющим собой смеси ...
... начало от одного конца полярона, так и в двух направлениях (с обоих концов полярона). Больше того, предполагают даже, что рекомбинация очень сходна по своему механизму с процессом транскрипции генетической информации (образования мРНК) в смысле определения конца гена, с которого начинается разделение ДНК, и что в рекомбинации также работает ген-оператор, сходный с оператором оперона. Анализ всех ...
... сходство хромосом по сравнению с другими клеточными органеллами к основным красителям. В течение последующих 10 – 15 лет большинством биологов было подтверждено, что именно хромосомы служат материальным носителем наследственности. Хромосомы особенно четко видны во время делений клеток, однако факт непрерывности их существования и в неделящихся ядрах сомнений не вызывает. Основная особенность ...
... на передний план наиболее важные (с точки зрения учебных целей и задач) характеристики изучаемых объектов и явлений природы[4]. 1.2 Методические приемы использования мультимедиа на уроках биологии. Преимущества мультимедийных технологий, по сравнению с традиционными, многообразны: наглядное представление материала, возможность эффективной проверки знаний, многообразие организационных форм ...
0 комментариев