Предмет, історія розвитку і завдання мікробіології
Мікробіологія – наука про мікроорганізми, про найдрібніші істоти біосфери, про форми живої матерії, що вони невидимі неозброєним оком.
Загальними ознаками мікроорганізмів є їх малий розмір, висока інтенсивність і пластичність метаболізму. Мікроорганізми перебувають скрізь, їх загальна біомаса перевершує масу рослин і тварин і складає 740 млрд. тонн.
Мікроорганізми вивчають за допомогою оптичного або електронного мікроскопу. Розмір вірусів коливається в діапазоні 16-200 нм, а розмір інших досягає 0,01- 50.
Наприкінці 20-го століття була розроблена класифікація, згідно якій, всі живі істоти біосфери поділяються на три надцарства:
- акаріоти (без’ядерні) – царство вірусів;
- прокаріоти (доядерні) – царства архебактерій, ціанобактерій та еубактерій;
- еукаріоти (ядерні) – царства рослин, тварин і грибів.
В історії розвитку розділяють 4 періоди:
Морфологічний. Історія мікробіології починається з кінця 17-го століття, коли голландський натураліст Антоній Ван Левенгук (1632-1723) відкрив світ мікроорганізмів. А.Левенгук створив перші мікроскопи, які збільшували предмети в 160-300 разів. Він з великою точність описав і зарисував зовнішній вигляд (морфологію) «живих звіряток». Цей період був періодом накопичення фактичного матеріалу.
Еколого-фізіологічний. З другої половини 19-го століття завдяки роботам великого французького мікробіолога Луї Пастера (1822-1895) було пов’язане створення мікробіології як науки.
Основні відкриття Л.Пастера та його внесок такі:
- встановив, що мікроорганізми є причинами інфекційних хвороб;
- аеробне і анаеробне бродіння це функції діяльності мікроорганізмів;
- запропонував метод пастеризації для лікування “хвороб” вин;
- розробив методи виготовлення живих вакцин проти сибірки і сказу;
- заклав основи науки імунології;
- встановив здатність мікроорганізмів засвоювати і перетворювати різноманітні хімічні сполуки.
Роберт Кох (1843-1910 розробив основи дезинфекції, метод виділення чистої культури мікроорганізмів на твердому поживному середовищі, способи забарвлення мікробів аніліновими барвниками, застосував імерсійну систему у мікроскопуванні та ін. Р.Кох відкрив збудників туберкульозу (паличка Коха) та холери.
Імунологічний. Видатний російський вчений І.І.Мечников (1845-1916) розробив теорію фагоцитарного імунітету, відкрив і теоретично обґрунтував антагонізм мікробів. І.І.Мечников є автором відомої теорії боротьби з передчасним старінням людського організму, для боротьби з яким він рекомендував вживати молочнокислі бактерії як антагоністи гнильних мікробів. М.Ф. Гамалія (1859-1949) відкрив станцію щеплення проти сказу. Д.Й. Івановський (1864-1920) став засновником вірусології. С.Н. Виноградський (1856-1953) відкрив хемосинтез. Видатний український мікробіолог Д.К.Заболотний (1866-1929)організував першу в світі кафедру епідеміології при Одеському медичному інституті. Багато зусиль віддав він вивченню чуми, холери, сифілісу, дифтерії, тифу тощо.
Молекулярно-генетичний. У цей період був вивчений механізм взаємодії облігатного паразиту і людини, рослини. Встановлено, що ДНК є носієм спадкової інформації. Відбулося створення біотехнології.
Основні напрямки сучасної мікробіології
Мікробіологія як наука поділяється на низку самостійних дисциплін – загальну, сільськогосподарську, медичну, ветеринарну, технічну, геологічну та космічну мікробіологію.
Загальна мікробіологія досліджує загальні закономірності життєдіяльності мікроорганізмів (анатомію, фізіологію, біохімію, генетику, екологію, систематику і еволюцію мікробів).
Сільськогосподарська мікробіологія розробляє методи та препарати для підвищення родючості грунту; фітопатогенів та розробляє методи боротьби з ними.
Медична мікробіологія вивчає мікроорганізми, що викликають захворювання у людей, та способи боротьби з цими захворюваннями.
Ветеринарна мікробіологія досліджує збудників захворювань у тварин та птахів і розробляє методи боротьби з ними.
Геологічна мікробіологія вивчає роль і значення мікробів у геологічних процесах.
Морська мікробіологія вивчає мікробів, які живуть в водних басейнах та їх вплив на водні екологічні системи.
Космічна мікробіологія досліджує вплив на мікроорганізми невагомості та інших чинників.
Технічна мікробіологія розробляє наукові основи використання мікроорганізмів у різних виробничих процесах. Винятково важливе значення мають мікробіологічні процеси в виробництві харчових продуктів.
Біотехнологія базується на інтегрованому використанні досягнень біохімії, мікробіології та інженерних наук. Завдяки застосуванню методів генної інженерії створюють трансгенні мікроорганізми, які є високопродуктивними продуцентами різноманітних біологічно активних речовин.
Систематика вивчає відношення між групами мікроорганізмів. За період становлення мікробіології в основу класифікації різноманітних організмів були закладені морфологічні, фізіологічні, біохімічні та інші ознаки.
Один з перших відомих поділів мікроорганізмів на групи базується на відношенні їх до субстрату життєдіяльності. За цим критерієм всі мікроорганізми поділяють на сапрофіти і патогенні.Сапрофіти – розвиваються на мертвих субстратах. Серед них розрізняють корисні – мікрофлора заквасок, молочнокислих продуктів, копчених ковбас, хліба, вина, пива. Але серед сапрофітів є велика кількість збудників псування продуктів, наприклад, гнильні бактерії.
Патогенні – паразити, які живуть на живих субстратах, використовують їх білки, жири, вуглеводи та інші складові частини. Їх поділяють на: зоопатогенні – паразити тварин ; фітопатогенні – паразити рослин. Таксон є основною категорією у ситематиці мікроорганізмів. У сучасній систематиці основною структурною одиницею є вид.
Порядок розташування таксонів: клон → вид → род → родина → порядок → клас → відділ → царство → надцарство.
Клон – мікроорганізм, який походить від клітини. Штам – чиста культура одного виду . Штами відрізняються другорядними властивостями. Наприклад відомі штами Clostridium botulinum A, B, C, D, E, F, I.
Групою вчених під керівництвом Бергі було видано визначник бактерій, який у 1997р. було перекладено російською мовою «Визначник бактерій Бергі».
Розподіл мікроорганізмів за таксонами здійснюється шляхом формування сукупностей споріднених організмів за категоріями:
- морфологічні ознаки. Форма клітин і характер їх розташування, розмір, забарвлення за Грамом, спороутворення, рухомість.
- культуральні ознаки. Вирощування мікроорганізмів на поживних середовищах зветься культивування, а їх нащадки - культурою. Змішані культури – це сукупність мікроорганізмів різних видів, які були виділені з біоценозу в природі (грунту, води, повітря). Чисті культури – нащадки одного виду, які виділені ізольовано на стерильному поживному середовищі. Культуральні ознаки – характеристика росту мікроорганізмів на середовищах.
- фізіологічні ознаки. Тип дихання, живлення, метаболізм, розмноження та ін.
- біохімічні властивості. Здатність мікроорганізмів продукувати визначені ферменти.
- резистентність до факторів середовища.
- токсиноутворення.
- антигенний склад. Антиген – речовина, яку виробляють патогенні мікроорганізми, коли потрапляють в організм хазяїна. При цьому відбуваються реакції імунітету, хазяїн продукує антитіла. Наприклад реакція аглютинації, яку записують:
RA = Ag+An
Досліджування всіх ознак у невідомих мікроорганізмів дозволяє за допомогою визначника Бергі ідентифікувати їх вид. Номенклатура – збірник правил найменування таксонів, список цих найменувань. В мікробіології застосовується бінарна термінологія – назва виду складається з двох : родова ( пишеться з великої літери) і видова ( пишеться з маленької літери). Родова – може носити прізвище вченого, показувати морфологію, наявність спор. Видова – може означати фізіологію, місце існування. Наприклад : Clostridium botulinum, Bacillus subtilis, Staphylococcus aureus.
Морфологічні типи бактеріальних клітин
Розміри бактерій коливаються від 0,2 до 10 мкм (мікрометрів). За формою клітини бактерії можна розділити на головні групи: сферичні або кулясті (коки), паличкоподібні (циліндричні) та спіралеподібні (звивисті), але відомі бактерії, які мають ниткоподібну, трикутну, зірчасту форми (див.рис.). Розміри та форма тіла бактерій можуть змінюватися під впливом різноманітних факторів зовнішнього середовища – під дією антибіотиків, дезинфікуючих засобів, при зміні умов культивування та ін.
Сферичні бактерії або коки (від грецького coccus - ягода, зерно) звичайно мають форму кулі, але бувають у формі боба чи ланцеподібні. Клітини можуть розташовуватися хаотично або утворювати скупчення, форма яких залежить від характеру їх поділу. За характером взаємо розташування коки поділяють на:
Мікрококи (від грецького micros - малий) діляться в одній площині, розташовуються хаотично, поодиноко. Є переважно сапрофітами. Проте у людей з імунодефіцит ним станом можуть викликати гнійно-септичні захворювання.
Диплококи (від грец. diplos - подвійний) також діляться в одній площині, але утворені пари клітин не розходяться. Типовими представниками є збудники епідемічного менінгіту, гонореї, та крупозної пневмонії.
Стрептококи (від грец. streptos – намисто, ланцюжок) діляться в одній площині, але зв’язок між клітинами зберігається, і утворюються ланцюжки різної довжини, які нагадують намисто. Серед стрептококів багато патогенних для людини, також є представники збудників молочно - кислого бродіння – Streptococcus lactis, Streptococcus cremosis.
Тетракоки (від грец. tetra - чотири) діляться в двох взаємо перпендикулярних площинах з утворенням тетрад. Вони переважно складають мікрофлору повітря, патогенні для людини види зустрічаються дуже рідко.
Сарцини (від лат. sarcina – поєдную, тюк) діляться в двох взаємно перпендикулярних площинах з утворенням “пакунків” з 8, 16, 32, 64 клітин, які розміщуються кількома ярусами. Особливо часто сарцини зустрічаються у повітрі і є пігментоутворюючими. Патогенних форм серед них немає.
Стафілококи (від грец. stafile - гроно) діляться в декількох площинах, клітини розташовуються у вигляді виноградного грона. Зустрічаються у повітрі, грунті, в організмі людини і тварини.
Паличкоподібні бактерії. Паличкова форма є самою численною і різноманітною групою бактерій. Клітинна стінка паличкоподібних бактерій являє собою тверду, нееластичну трубку. Тому вони ростуть переважно в довжину, а коливання товщини відносно невеликі (переважно в межах 0,5-1 мкм). По довжині паличкові поділяють на дрібні (до 1 мкм), середні (2-4 мкм), великі (більш ніж 4 мкм). По товщині паличкові бактерії поділяють на тонкі () до 1 мкм і товсті (>1 мкм). Форма кінців у паличкових буває закруглена, загострена, обрубана.
По здатності до спороутворення паличкоподібні форми бактерій підрозділяються на три роди: р.Bacterium (бактерії), р.Bacillus (бацили), p.Clostridium (клостридії).
p.Bacterium – це паличкоподібні бактерії, що, як правило, не утворюють спор. Типовим представником є кишкова паличка (Escherichia coli), яка завжди міститься в кишечнику людини та тварин.
p.Bacillus – це споротворні паличкоподібні бактерії. Діаметр спор не перевищує у них поперечного розтину палички тому спороутворення у бацил не призводить до зміни форми клітини. За типом дихання бацили, як правило, відносяться до аеробів (потребують кисень для свого розвитку). Прикладом сапрофітних форм бацил є Bacillus subtilis (“сінна” паличка), Bacillus licheniformis (“картопляна” паличка).
p.Clostridium – це споротворні паличкоподібні бактерії, у яких спора за своїм розміром перевищує поперечний розтин палички. При цьому змінюється зовнішня форма палички і вона може нагадувати веретено або ракетку. За типом дихання відносяться. Як правило, до облігатних анаеробів.
За взаємним розташуванням паличкові розподіляються на три підгрупи:
а) поодинокі палички – розміщуються хаотично. Така форма взаємного розташування клітин характерна для переважної більшості паличкових.
б) диплобактерії та диплобацили – клітини бактерій або бацил розташовані попарно.
в) стрептобактерії та стрептобацили – клітини бактерій або бацил утворюють ланцюжки різної довжини.
Звивисті бактерії. Вони як правило грамнегативні, рухомі, більшість належить до сапрофітів. Але зустрічаються і патогенні форми – збудники інфекційних хвороб. Звивисті бактерії відрізняються між собою кількістю завитків та товщиною тільця.
В залежності від цього відрізняють:
- вібріони. Клітини мають один завиток і товсте тільце. Представником є холерний вібріон – збудник холери, водні вібріони.
- спірили. Клітини мають від 4 до 6 завитків спіралі і товсте тільце.
- спірохети. Клітини мають від 6 до 15 і більше завитків спіралі та тонке тільце. Довжина їх може бути великою (300-500 мкм). Представником є збудник венеричного захворювання – сифілісу (Treponema pallidum).
Ультраструктура бактеріальної клітини
В 1937 р. Е.Шаттоном було запропоновано поділяти всі живі істоти за будовою клітин на прокаріоти та еукаріоти.
Згідно з цією класифікацією до прокаріотів (грец. procaryotae- доядерні) відносяться бактерії, до еукаріотів ( грец. eucaryotae- ядерні) – найпростіші, гриби, водорості, рослини та тварини.
Основні відмінності прокаріотів та еукаріотів полягають в тому, що прокаріоти мають тільки одну внутрішню порожнину, яка утворена цитоплазматичною мембраною (ЦПМ), що розміщена під клітинною стінкою. Їхній ядерний апарат (нуклеоїд) не відособлений від цитоплазми. Він складається з однієї хромосоми, яка має вигляд замкненої в кільце молекули ДНК, та розміщена безпосередньо в цитоплазмі. Рибосоми прокаріотів мають незначні розміри та розсіяні в цитоплазмі.
В клітинах еукаріотів є вторинні порожнини, утворені ЦПМ – ядро та органели. Ядро містить ДНК у вигляді однієї або більше хромосом та ядерце. Органели представлені мітохондріями, хлоропластами, лізосомами, ендоплазматичною сіткою, апаратом Гольджи та інш. структурами. Рибосоми еукаріотів більші за рибосоми прокаріотів та розміщені переважно на поверхні ендоплазматичної сітки.
Анатомія бактерій вивчає за допомогою електронного мікроскопу внутрішню будову клітин. Клітина – біологічна система, у якій кожній структурі належить певна функція. Всі структури взаємопов’язані та взаємообумовлені.
Основні структури клітин бактерій:
- ядерний матеріал – нуклеоїд
- клітинна стінка
- цитоплазматична мембрана (ЦПМ)
- цитоплазма з органоїдами і включеннями
- структурні елементи клітини (рибосоми, мезосоми, плазміди,
внутрішньоклітинні мембранні утворення, включення )
Додаткові структури:
- капсула
- слиз
- пілі
- джгутики
- фімбрії
Ядерний матеріал клітин бактерій
У прокаріотів ядерний матеріал розміщений у центральній частині цитоплазми. Нуклеоїд бактерій на відміну від еукаріотів не має мембрани і складається з ДНК, яка замкнута у кільце. Приймає участь в передачі спадкової інформації. Без нього клітина нежиттєздатна.
Багато бактерій разом з нуклеоїдом мають позахромосомні передавачі спадкової інформації. Їх називають плазміди або епісоми . За будовою це нуклеїнова кислота.
Клітинна стінка
Клітинна стінка – обов’язковий структурний елемент бактеріальної клітини. Клітинна стінка надає бактеріям певну форму. Вона є пружною, але разом з цим еластичною і може вигинатися. Клітинна стінка виконує роль механічного бар’єру між зовнішнім середовищем та протопластом.
Вона захищає вміст клітини від впливу довкілля. В нормальних умовах концентрація солей в клітині вища, ніж у навколишньому середовищі, тому між ними існує велика різниця в осмотичному тиску. Клітинна стінка захищає клітину від проникнення до неї надлишку води.
Таким чином функції клітинної стінки слідуючі:
- захист від шкідливих факторів
- транспорт речовин в процесі живлення
- участь у формуванні капсул і антигенів
Товщина клітинної стінки знаходиться в межах від 10 до 80 нм, на її долю припадає від 5% до 50% сухої речовини клітини. Хімічний склад і будова клітинної стінки є постійними для певного виду бактерій. Клітинна стінка відповідальна за сприйняття забарвлення за Грамом.
До складу клітинної стінки бактерій входить мурен, він є гетерополімером, який побудований з N-ацетилглюкозамін, N-ацетилмурамової кислоти. Вони утворюють ланцюги, які пов’язані пептидними зв’язками і утворюють муреїнову сітку. У грампозитивних бактерій клітинна стінка складається з багатошарового мурену (до 9% її маси), тейхоєвих кислот, лізіну і невеликої кількості ліпідів, полісахаридів, білків. У грамнегативних бактерій клітинна стінка значно тонша, вона містить одношаровий мурен (до 10%), ліпополісахариди, ліпопротеїди, фосфоліпіди, діамінопімелінову кислоту.
Деякі мікроорганізми на поверхні клітин утворюють слизовий шар або капсулу. Хімічний склад шару полісахариди, поліпептиди. Слиз можуть утворювати бактерії Leuconostoc, викликає псування фруктових сиропів.
Функції слизу і капсул – захисна. У патогенів капсула надає стійкість і підвищує вірулентність – ступінь патогенності.
Рухомість бактерій
Рухомість бактерій пов’язана з наявністю джгутиків. Тип джгути кування є систематико – діагностичною ознакою і генетично запрограмований. Кількість і локалізація джгутиків, їх розміри – характерні визначеному виду.
В залежності від розташування джгутиків рухливі бактерії поділяють на наступні групи: а) монотрихи – мають один джгутик на кінці
б) амфітрити – мають два полярно розташований джгутики
в) лофотрихи – мають пучок джгутиків на одному кінці
г) амфілофотрихи – мають два пучки полярно розташованих джгутиків
д) перитрихи – мають хаотично розташовані джгутики по всій поверхні клітини
Швидкість руху бактерії залежить від кількості та характеру розташування джгутиків, від віку і факторів зовнішнього середовища.
Джгутик має вигляд спіральної нитки товщиною 10-20 нм і довжиною до 10 мкм, їх хімічна природа – білок флагелін. Рухомість бактерій визначають методами «висячої» і «роздавленої» краплин. Бактерії рухаються хаотично зі швидкістю до 100 мкм/сек , але здатні до таксисів – спрямованих рухів. Відомі хемо- , аеро- , фототаксис, вони залежать від концентрації хімічних речовин, кисню, ступені освітлення відповідно.
На поверхні клітинної стінки деяких мікроорганізмів без джгутиків розміщуються або фімбрії, або пілі – довгі, прямі нитки діаметром 5 нм, довжиною 12 мкм. Їх кількість коливається і може досягати декількох тисяч. Вони бувають загального типу для прикріплення і «статевого» в процесі кон’юктації. Хімічна природа цих структур – білок пілін. Фімбрії також мають вигляд довгих тонких прямих ниток, а їх хімічна природа – вуглеводозв’язувальні білки лектини.
Цитоплазматична мембрана
Під клітинною стінкою розташована цитоплазматична мембрана (ЦПМ), яка є обов’язковим структурним елементом клітини і оточує цитоплазму. Бактеріальна клітина втрачає життєздатність при порушенні її цілісності. На долю ЦПМ припадає всього 8-15 % сухої речовини клітини. Загальна товщина приблизно 5-10 нм. У більшості прокаріотів ЦПМ єдина мембрана.
ЦПМ утворена двома шарами ліпідів (ліпідний біслой), в які включені білки, а зовні розташовані периферичні білки. За хімічним складом вона містить 50-75% білків, 15-45% ліпідів та незначну кількість вуглеводів.
ЦПМ грає найважливішу роль в метаболізмі бактеріальної клітини. Вона є осмотичним бар’єром і регулює осмотичний тиск в клітині, контролює транспорт речовин в клітину та виведення метаболітів назовні. У бактерій ЦПМ виконує енергетичну функцію. ЦПМ бактерій – структура, де відбуваються окисно-відновні процеси. ЦПМ – енергетичні станції з ферментами окислювального фосфорилювання. ЦПМ приймає участь в поділі нуклеоїду, оскільки молекула ДНК при цьому процесі прикріплюється до неї. Вона бере участь у синтезі компонентів клітинної стінки та капсули.
Цитоплазма
Цитоплазма являє собою колоїдну систему, яка складається з двох фракцій: цитозоль та структурні елементи.
Цитозоль – це фракція цитоплазми, яка має гомогенну консистенцію. У молодих вегетативних клітин міститься 85% води. В воді утворюється колоїдна система з білків, нуклеотидів, БАР, вітамінів, осмотично-активних речовин, вуглеводів, мінеральних елементів, токсинів. Друга фракція включає різноманітні структурні елементи клітини: генетичний апарат, плазмідм, рибосоми, мезосоми, включення, внутрішньоклітинні структури, вакуолі.
Рибосоми – це органоїди, мають 10-20 нм в діаметрі і складаються з РНК (60%) та білка (40%).Кожна бактерія містить від 5000 до 50000 рибосом, які виконують важливу фізіологічні функцію – синтез білків. Під час активного синтезу білків рибосоми утворюють специфічні ланцюги - полісоми. Синтез білка йде в 2 етапи: за один етап нова молекула білка збільшується на одну молекулу амінокислоти.
Багато бактерій разом з хромосомною ДНК мають в цитоплазмі позахромосомні кільцеві або лінійні ДНК дуже невеликого розміру, які називають плазміди. В плазмідах закодована важлива інформація, що визначає життєдіяльність клітини в певних умовах, в тому числі хвороботворність, структуру ферментів, які дозволяють клітині знешкоджувати деякі антибіотики та інше.
Мезосоми – це тільця розміром 100 А, вони утворюються з ЦПМ. У ЦПМ існують інвагінації - складки. Вони мають форму бульбашок, можуть нагадувати сітку, можуть бути трубчасті, пластинчасті. Там локалізовані окисно-відновні ферменти, тобто енергетичний обмін. Поруч з цим мезосоми – місце прикріплення плазмід, синтезу ферментів.
Внутрішньомембранні структури виконують різноманітні функції. У прокаріот вони забезпечують дихання, захист від УФ променів, фотосинтез, окисно-відновні процеси. В них містяться бактерії, хлорофіл, каротин та інші пігменти.
Включення – гранули різної форми та розмірів або краплі. Наявність їх в клітині не можна вважати за постійну ознаку мікроорганізму, оскільки включення зв’язані з фізичними та хімічними умовами середовища існування та можуть розглядатися як запасні джерела енергії та поживних речовин.
Будова спори
екзоспоріум
зовнішня оболонка спори
внутрішня оболонка спори
корт екс (кора)
клітинна стінка зародку
цитоплазматична мембрана
цитоплазма з ядерною речовиною
Рис. Будова зрілої ендоспори
Спори бактерій – ендоспори – внутрішньоклітинні утворення круглої або овальної форми. При забарвленні бактерій за Грамом клітина – синя, а спора прозора. Продукція спор – стадія циклу розвитку паличкоподібних грам позитивних Bacillus та Clostridium (виняток Lactobacillus), яка виробилась в процесі еволюції в боротьбі за збереження виду. Функція спор бактерій – захист. Спори знаходяться в стані анабіозу – не розвиваються, не розмножуються, але зберігають життєздатність. При попаданні в благо приємні умови одна спора проростає в одну вегетативну клітину.
Спори утворюються в несприятливих умовах, під впливом фізичних та хімічних факторів: низька вологість, температура, висока концентрація речовин, радіація, УФ промені. Процес спороутворення йде від 18 до 72 годин і відбувається по стадіям. Спочатку утворюється спорогенна зона і проспора, потім йде дозрівання спори. На 1 етапі втрачається вода, розпадаються материнські білки, синтезується специфічна лише для спори діпіколінова кислота, активно поглинається кальцій, який з нею дає халатні комплекси. На 2 етапі йдуть формування багатошарової оболонки і всіх структур. Спора відрізняється від вегетативної клітини підвищеною стійкістю до несприятливих умов.
Властивості спор:
- термостійкість. В той час, коли вегетативні клітини гинуть при 80 °С за 10 хвилин, спори гинуть при автоклаву ванні протягом 24 хвилин при температурі 120°С при тиску 1 атм (1,61∙10³Па). У висушеному стані вони гинуть при нагріванні до 150-160°С протягом декількох годин.
Спори Clostridium botulinum витримують кип’ятіння протягом 5-6 годин. Спори окремих видів бактерій відрізняються особливою термостійкістю. Спори Clostridium botulinum можуть складати залишкову мікрофлору стерилізованих консервів, також спори Clostridium botulinum являються еталонною тест – культурою при науковому обґрунтуванні режимів стерилізації.
Терморезистентність спор обумовлена низьким вмістом води (до 40%, вона знаходиться у зв’язаному стані) та вмістом дипіколінату кальцію.
- хімічна стійкість спор обумовлена непроникністю багатошарової оболонки.
- стійкість до радіації, УФ-променів пов’язана зі значною концентрацією сірковмісних амінокислот (цисті, цистеїн).
В зовнішньому середовищі спори можуть зберігатись тисячі років.
Розрізняють декілька видів типів спороутворення (див. рис.). Якщо при утворенні спори в центрі клітини форма її не змінюється, то такий тип спороутворення називається бацилярним, він властивий представникам роду Bacillus. Коли клітина в середині потовщується та набуває вигляду веретина, то такий тип спороутворення називається клостридіальним (він характерний для збудника гниття білків Clostridium sporogenes). Іноді спора утворюється ближче до кінця клітини і тоді клітина набуває вигляду тенісної ракетки – це плектридіальний тип спороутворення (він притаманний збуднику ботулізму Clostridium botulinum). Клостридіальний та пектридіальний типи властиві бактеріям роду Clostridium.
Мікроміцети
Мікроміцети – це численна та різноманітна група, яка включає до 100 тисяч видів. Вони дуже поширені у природі і зустрічаються майже в усіх кліматичних зонах. Найбільш розповсюджені у грунтах, а також у водоймищах. Утворюють на поверхні субстратів цвіль різного кольору, тому їх називають плісеневими грибами.
За типом дихання – аероби і розвиваються переважно на поверхні субстрату. Але є і мікроаерофіли, які розвиваються при невисоких концентраціях кисню (<5%) і здатні проростати в середину субстрату.
За типом живлення гриби – хемогетеротрофи. Джерелом вуглецю для них можуть бути вуглеводи, органічні кислоти, білки.
Джерелом азоту для плісеневих грибів є амінокислоти, білки, а також неорганічні речовини.
Таким чином, субстратом для розвитку грибів можуть бути як харчові продукти, сировина і корми, так і технічні матеріали (целюлоза, резина, фарби).
Псування продуктів, що викликають плісеневі гриби, називається пліснявінням.
Більшість грибів є мезофілами і віддають перевагу середнім температурам 16-27°С. Температурний оптимум у грибів нижчий, ніж у бактерій, є холодолюбиві види, які відносяться до психрофілів. Найбільш холодостійкі види – Cladosporium gerbarum, Thamnidium elegans. Гриби здатні розвиватися при вологості повітря 70-100%. Серед них є і ксерофіти, які розвиваються при вологості до 65%.Вони псують солодку продукцію з низьким вмістом вологи (мармелад, джем, пастилу).
Інтоксикації грибкової природи називають мікотоксикозами, а метаболіти грибів, які мають токсичні властивості, називають мікотоксинами. Відомо більш двохсот індивідуальних мікотоксинів. Найбільш повно вивчена хімічна природа афлатоксинів, зеараленону, трихотеценів, охратоксинів, стеригматоцистину.
Більшість мікотоксинів термостійкі, канцерогенні, мають кумулятивні властивості. Мікотоксини можуть зберігатися в продуктах навіть при стерилізації.
Для запобігання ураження плісеневими грибами загальноприйняті технології зберігання сировини доповняють обробкою спеціальними хімічними препаратами – плісеневими інгібіторами, які містять органічні кислоти та їх солі.
Пряме сонячне світло гальмує розвиток плісеней, але чергування світла і темряви стимулює ріст і спороутворення. Багато видів грибів, що містять пігменти, є стійкими до УФ світла і радіації.
Плісеневі гриби мають і позитивне значення для харчової промисловості. Деякі гриби використовують у заквасках для виробництва кисломолочних продуктів і сирів, для одержання ряду органічних кислот (лимонної, щавлевої, яблучної), антибіотиків, вітамінів, ферментів.
Плісеневі гриби можуть синтезувати різні вторинні метаболіти, багато з яких володіють біологічною активністю. Метаболіти грибів, які здатні викликати токсичну дію у людей і тварин, наз. мікотоксинами.
Морфологія та будова клітин мікроміцетів
Вегетативне тіло грибів називається міцелієм або грибницею. Міцелій являє собою систему розгалужених ниток-гіфів, що складаються з довгих клітин, які розміщені в один ряд. Міцелій буває двох типів: септований і несептований. Септи являють собою поперечні перегородки, які поділяють міцелій на ряд окремих клітин.
Септи мають центральну пору, через яку з клітини в клітину вільно перетікають цитоплазма і ядра. Більша частина гіфів розвивається над поверхнею субстрату і називається повітряним або поверхневим міцелієм. Саме тут розташовані органи розмноження.Гриби мають еукаріотну будову клітини, яка подібна до будови рослинних клітин.
Зовні клітина грибів покрита багатошаровою стінкою, яка на 80-90% складається з полісахаридів (хітин, целюлоза). Під стінкою розташована ЦПМ, яка оточує цитоплазму. В цитоплазмі містяться ядра (від 1 до 30). Ядро оточене власною ядерною мембраною з порами, містить ядерце і хромосоми. В ядерці відбувається синтез попередників рибосом, які транспортуються через пори ядра в цитоплазму. В цитоплазмі містяться 30-50 тисяч рибосом. В клітинах грибів є система внутрішньо - клітинних елементарних мембран.
Клітини грибів містять органоїди: ядро, мітохондрії, ендоплазматичну сітку, апарат Гольджи, лізосоми.
Мітохондрії – багатокамерні трубочки з еластичними стінками. Вони є енергетичним центром клітини.
Ендоплазматична сітка - мембранна система, в якій розташовані ферменти, що відповідають за біосинтез вуглеводів, ліпідів і транспорт речовин всередині клітини.
Апарат Гольджі – здійснює транспортування речовин, що синтезуються в ендоплазматичній сітці, і вилучення продуктів обміну.
Лізосоми – розщеплюють білки, ліпіди, полісахариди до простих сполук. У випадку пошкодження лізосом ферменти виходять в цитоплазму і здійснюють лізис (розчинення усіх органоїдів).
Способи розмноження мікроміцетів
Міцелі альні гриби розмножуються вегетативно та за допомогою спор.
Вегетативне розмноження відбувається за допомогою шматочків гіфів та міцелію. Будь-яка частина міцелію здатна до росту. Шматочки міцелію відриваються від гриба та розносяться вітром, попадають на відповідний субстрат, проростають і утворюють новий організм.
Спори в еукаріотичних організмах виконують подвійну функцію – розмноження та збереження виду. Спороутворення може відбуватися статевим і безстатевим шляхом.
Безстатеве спороутворення зв’язане з утворенням спеціалізованих репродуктивних органів без попереднього злиття клітин. Безстатеве розмноження нижчих грибів здійснюється внутрішніми спорами або ендоспорами, вищих грибів – зовнішніми спорами або екзоспорами.
Ендоспори утворюються в середині особливої клітини, яка наз. спорангієм . При дозріванні оболонка під впливом ферментів грибів і вологи розпливається і спори висипаються у зовнішнє середовище.
Екзоспори прийнято називати конідіями або конідієспорами, а плодоносний гіф – конідієносцем. Конідії розповсюджуються повітряними потоками і в сприятливих умовах проростають, утворюючи новий міцелій.
Статеве розмноження грибів обов’язково включає злиття двох ядер. У нижчих грибів спочатку відбувається злиття двох багатоядерних гіфів міцелію. Потім відбувається попарне злиття ядер. Процес закінчується утворенням зігоспори, яка згодом проростає і утворює органи плодоношення.
У вищих грибів в залежності від класу розмноження відбувається по різному.
В аскоміцетів – злиття ядер і вмісту двох клітин різних гіфів, потім відбувається ділення ядра.
Утворення зигоспори. Утворення аскоспор
Навколо нових ядер концентрується цитоплазма і утворюється спорова оболонка. Материнська клітина покривається товстою оболонкою і перетворюється в аск (сумку). Всередині може бути від 2 до 8 аскоспор.
Зверху сумка покривається переплетенням гіфів. При цьому відбувається утворення плодового тіла.
Систематика мікроміцетів
Мікроміцети поділяють на шість класів: хітридіоміцети, ооміцети, зигоміцети, аскоміцети, базидіоміцети та дейтероміцети.
Хітридіоміцети (Chytridiomycetes). У них міцелій відсітній або слабо розвинутий, тіло гриба являє собою голу цитоплазму. Безстатеве розмноження відбувається за допомогою рухомих зооспор, які утворюються в зооспорангіях. Багато представників цього класу є водними організмами, які розвиваються на відмерлих водоростях або паразитують на них. Деякі існують в грунті, інші відомі як паразити рослин.
Ооміцети (Oomycetes) являють групу організмів з характерним статевим процесом – оогамією та рухливими зооспорами з двома джутиками, які утворюються при безстатевому спороутворенні. Більшість ооміцетів є облігатними паразитами, що існують на рослині-хазяїні. Представники родів Pithium, Phytophthora викликають хвороби с/г рослин.
Зигоміцети (Zygomycetes) включають декілька порядків, найбільш поширений є мукоральний (Mucorales), що містить 14 родин. Серед мукорових грибів можна відмітити роди Mucor, Thamnidium, Rhizopus.
Гриби роду Mucor («головчаста плісень») можуть розвиватися на продуктах рослинного походження.
а- спори
б- спорангій
в- спорангієносець
Вони часто псують вологе зерно. Деякі види здатні викликати спиртове та молочно-кисле бродіння. У промисловості мукорові гриби використовуються для виробництва оргенічних кислот, ферментів, каротиноїдів та стеринів. Можуть рости на стінах вологих проміщень у вигляді сірого пухнастого нальоту.
Гриби роду Rhizopus відрізняються тим, що їх спорангієносці ростуть групами у вигляді пучків по декілька штук зі сплетення гіфів, що називається вузлом. Спорангієносці зверху мають потовщення – апофізу. Гриб утворює ризоїди, які є органами прикріплення гриба до субстрату і нагадують коріння рослин.
З їх допомогою гриб отримує поживні речовини з субстрату. Органами розселення гриба по субстрату є столони. Гриби цього роду вражають ягоди, коренеплоди, бульби, викликаючи «м’яку» гнилизну. Часто зустрічається на харчових продуктах – хлібі, рисі, борошні, плодах та овочах.
Гриби роду Thamnidium відрізняються тим, що у головного спорангієносця є бокові відгалуження виделкоподібної форми, а на їх кінцях розташовані невеликі спорангії – спорангіоли.. Вони містять одну або невелику кількість спор.
Вид Thamnidium elegans є найбільш холодостійким, тобто психрофітом і здатний псувати продукти навіть в холодильних камерах. Викликає глибинне псування продуктів, синтезує мікотоксин, тому м’ясо і м’ясні вироби уражені Thamnidium elegans не придатні для використання.
Аскоміцети (Ascomycetes) або сумчасті гриби. До аскоміцетів, які мають особливе значення в житті людини, відносяться представники родів Aspergillus і Penicillium. Ці гриби – типові мешканці грунту, а також виявляють в повітрі, на зерні, плодах, овочах, харчових продуктах.Серед грибів цих родів відомі продуценти таких антибіотиків як пеніцилін, цитрин, фумагілін та ін.
Деякі види синтезують мікотоксини.
Рід Aspergillus (лієчка плісень) характеризується розгалуженим багатоклітинним септованим міцелієм. Конідієносці несептовані. На вершині відростають китички. На китичках ланцюжками розташовані конідії (див.рис.а), які часто забарвлені.
Деякі види аспергилів використовуються у промисловості – для отримання лимонної кислоти, при виробництві ферментних препаратів, для освітлення соків.
Серед аспергилів є збудники грибних захворювань – мікозів.
Гриби роду Penicillium мають багатоклітинний, розгалужений септований міцелій. Конідієносці також септовані (див.рис.б)
Конідії розносяться потоками повітря і проростають. Певні види Penicillium використовують для отримання ферментних препаратів.
Базидіоміцети (Basidiomycetes). Орган спороношення – базидія.
До них належать сажкові та іржасті гриби. Базидіоміцетам є шкідник деревини, їстівні гриби, а також різноманітні сапрофіти, що беруть участь у розкладі органічних залишків.
Дейтероміцети (Deuteromycetes) мають добре розвинений септований багатоклітинний міцелій. Розмножуються безстатевим шляхом – конідіями. Багато представників даного порядку – Cladosporium, Alternaria, Fusarium, Botrytis.
Гриби роду Fusarium мають розгалужений багатоклітинний міцелій, розмножуються конідіями і поділені септами на клітини.
1- Fusarium
2-Botrytis
3-Alternaria
4-Cladosporium
5- Phoma
6- Catenularia
а- конідії
б- конідієносець
Конідієносці відсутні, конідії формуються на кінцях гіфів міцелію. Fusarium вражає картоплю, томати, буряк, цибулю, часник, злакові культури.
При вживанні хліба із зерна, враженого цими грибами, виникає захворювання, що наз. «п’яна хвороба». При цьому спостерігається слабкість, головний біль, блювання, розлад зору, біль у шлунку.
Рід Botrytis має багатоклітинний міцелій, конідієносці розгалужені, септовані. Вид Botrytis є збудником сірої гнилизни овочів та ягід. Міцелій гриба утворює на поверхні вражених овочів і ягід павутиноподібний наліт білого або сірого кольору. Конідії гриба мають сірувато-зелений колір, що і дало назву псуванню.
Рід Alternaria має конідії гранатоподібної форми, що мають декілька поперечних і повздовжних перегородок. Конідії мають темний колір, розташовані поодиноко або сполучені у ланцюжки, сидять на слаборозвинутих конідієносцях.
Різні види Alternaria широко розповсюджені у грунтах і викликають у с/г рослин чорну гнилизну (альтернаріоз).
Рід Cladosporium має прямі слаборозвинуті конідієносці. На них розвиваються ланцюжки конідій різної форми і розмірів. Зустрічаються кулясті, овальні, циліндричні конідії; конідії з однією або декількома септами, інколи без септ. Певні види добре розвиваються при низьких температурах. Цей гриб виявляється також на сирі, ковбасах, яйцях, маслі.
Принципи класифікації, морфологія та будова клітин дріжджів
Дріжджі – одноклітинні гриби, що не утворюють справжнього міцелію. Дуже поширені у природі, переносяться дощем, вітром, комахами і найбільш часто зустрічаються на рослинах, де є цукристі речовини. Дріжджі грампозитивні нерухомі організми. За типом живлення – хемогетеротрофи, за типом дихання – факультативні анаероби. Але є невелика група дріжджів, які розвиваються на поверхні субстратів і за типом дихання є аеробами.
Клітини дріжджів мають відносно великі розміри (в середньому 2-12 мкм). Дріжджі можуть суттєво змінювати свої форми та розміри.
Клітини дріжджів можуть мати різноманітну форму. Зустрічаються кулясті, овальні, еліптичні, циліндричні, лимоноподібні та інші форми.
За будовою клітини дріжджі відносяться до еукаріот. Дріжджова клітина має клітинну стінку. Під клітинною стінкою розміщена цитоплазматична мембрана, що охоплює цитоплазму клітини, в якій містяться органели та включення. Рибосоми у дріжджів розміщені в цитоплазмі і на зовнішньому боці ядерної мембрани. Ядро дріжджів оточене двошаровою мембраною і містить ДНК у вигляді хромосом.
Клітини дріжджів на 75% складаються з води. Суха речовина містить 90-95% органічних сполук – білків (30-75%), вуглеводів (25-50%), ліпідів (2-5%). Деякі дріжджі здатні синтезувати пігменти каротиноїди (Rhodotorula, Rhodosporidium).
Способи розмноження дріжджів
Дріжджі розмножуються вегетативно і за допомогою спороутворення, яке буває статевим і безстатевим. До вегетативних способів розмноження відносяться брунькування, поділ, брунькування поділом. При будь-якому вегетативному способі розмноження попередньо відбувається ділення ядра.
Вегетативне розмноження. Брунькування – найбільш розповсюджений спосіб розмноження. Воно характерне для дріжджів, що мають кулясту і овальну форму.
Розмноження поділом відбувається внаслідок утворення в дріжджовій клітині однієї або декількох поперечних перегородок – септ. Цей спосіб розмноження характерний для дріжджів циліндричної форми.
Брунькування поділом характеризується тим, що утворення дочірніх клітин починається з брунькування, а закінчується утворенням добре помітної септи в районі перешийка. Такий спосіб характерний для дріжджів лимоноподібної форми.
Спороутворення спостерігається у дріжджів при несприятливих умовах і відбувається після багаторазового розмноження шляхом брунькування. Оптимальна температура спороутворення 20-30°С.
Безстатеве спороутворення відбувається без попереднього злиття дріжджових клітин. Ядро ділиться на декілька частин – відповідно числу майбутніх спор. Ці спори знаходяться всередині материнської дріжджової клітини. Клітина виконує роль аска, тому наз. аскоспорами. При дозріванні аскоспор оболонка клітини розривається і спори попадають у зовнішнє середовище і проростають у вегетативні клітини.
Статеве розмноження дріжджів відбувається шляхом копуляції. Внаслідок копуляції можуть утворюватися аскоспори (у аскоміцетів) або екзогенні спори спори дії (у базидіоміцетів).
В цілому, спороутворення у дріжджів має подвійну функцію: розмноження і формування стійких видів. Спори дріжджів менш стійкі, ніж спори бактерій. Вони витримують температуру тільки на 10°С більшу, ніж вегетативні клітини дріжджів.
Характеристика найважливіших для промисловості родів дріжджів
Найбільше значення мають «культурні» дріжджі роду Saccharomyces, які викликають енергійне спиртове бродіння. У виробництві етилового спирту, пива, квасу, при приготуванні дріжджового тіста використовуються дріжджі Saccharomyces cerevisiae. Ці дріжджі мають овальну форму.
У хлібопеченні використовують раси дріжджів Saccharomyces cerevisiae, що активно виділяють вуглекислий газ, який сприяє доброму розпушенню і підніманню тіста.
Дріжджі поділяються на верхові та низові.
Верхові дріжджі здійснюють процес бродіння при відносно високих температурах (20-28°С). Процес бродіння відбувається бурхливо і забезпечує високий вихід спирту.
У виноробстві та пивоварстві використовують низові дріжджі. Низові дріжджі здійснюють процес бродіння при відносно низьких температурах (5-10°С). Спиртове бродіння протікає більш спокійно. Вони утворюють порівняно мало піни і каламуті, формують компактний осад у вигляді пластівців. Найбільш поширені у виноробстві винні дріжджі Saccharomyces vini, які мають еліптичну фору. До «диких» дріжджів відносяться дріжджі роду Zygosaccharomyces rautensteinii. Ці дріжджі осмофільні. Розвиваються при високих концентраціях цукру і солі (60-80%). Можуть псувати мед, сироп, варення, джем, згущене молоко з цукром, засолене м’ясо та рибу. Розмножуються статевим шляхом, викликають дуже повільне бродіння і в результаті накопичують до 10% спирту.
Дріжджі роду Schizosaccharomyces pombe є енергійними збудниками спиртового бродіння, але воно відбувається повільно. Можуть зброджувати глюкозу, сахарозу, мальтозу і декстрини, також яблучну кислоту в спирт і вуглекислий газ. Мають циліндричну форму із закругленими кінцями. Розмножування відбувається поділом. Використовують у виробництві рому і деяких сортів пива.
Дріжджі роду Saccharomycodes мають великі розміри, розмножуються брунькуванням поділом. Молоді клітини овальної форми, а дозрілі – лимоноподібної. Здійснюють спиртове бродіння з утворенням до 10% спирту. Це шкідники виноробства, які викликають помутніння вин, утворюють оцтово-кислий ефір, що надає вину гострий неприємний прокислий запах.
Рід Candida має клітини овальної і циліндричної форми. Вони є аспарогенні дріжджі. Рід Candida розмножується брунькуванням. Не викликають спиртового бродіння і тому є шкідниками бродильного виробництва.
Деякі види цього роду завдають шкоди дріжджовому виробництву, тому що якість хлібопекарських дріжджів знижується. Різні види роду Candida використовуються для отримання кормового білка та білково-вітамінних концентратів. З 500кг дріжджів можна отримати 50т білка. Існують патогенні види, які викликають кандидози, тобто уражають слизові оболонки рота, а інколи шкіри.
Дріжджі роду Torulopsis викликають спиртове бродіння і тому використовуються у виробництві кефіру і кумису. Мають кулясту, інколи овальну форму. Розмноження – численне брунькування. У виноробстві вважаються шкідниками, тому що утворюють слиз. Деякі дріжджі цього роду використовують для промислового одержання кормового білка.
Дріжджі роду Rhodotorulа відіграють певну роль у виробництві молочнокислих продуктів. Дріжджі можуть розмножуватися у молоці і мають здатність зброджувати лактозу. Але розвиток дріжджів в молоці більш повільний, ніж молочнокислих бактерій. Молочнокислі продукти, що виробляються з використанням заквасок дріжджів, мають лікувально-профілактичні властивості, тому що дріжджі синтезують антибіотичні речовини, активні проти патогенних штамів кишкової палички і збудника туберкульозу.
Віруси
Відмінні ознаки вірусів, будова віріонів, морфологія вірусів
В 1880 році Пастер застосував вакцини для лікування сказу, але про вірусну природу цієї інфекції, в той час було ще невідомо. Основоположником вірусології зїявися Івановський, він в 1892 р. відкрив збудника тютюнової мозаїки, якого не затримував бактеріальний фільтр.
Віруси (від лат. virus - отрута) –особлива форма життя, для якої відмінними ознаками, за якими вона відрізняється від інших живих істот є те, що віруси не мають клітинної будови, містять тільки один вид нуклеїнової кислоти та не здатні репродуктувати поза живою клітиною. Віруси – облігатні паразити.
Окрема зріла вірусна частка одержала назву віріон. Віріон складається з однієї молекули нуклеїнової кислоти та білкової оболонки. Оболонку віріонів називають капсидом (від сарsa – вмістилище). Капсиди утворюються з білкових субодиниць (поліпептидів), які називаються капсомерами. Їх кількість та метод укладки є постійним для кожного виду вірусів і використовується як таксономічна ознака.
Капсид віконує захисні йункцї і має досить високу стабільність до різних факторів внутрішньоклітинного середовища і особливо, до дії протеолітичних ферментів. Друга функція капсида полягає у забезпеченні адсорбції вірусів лише на тих клітинах, де може відбуватись їх розмноження.
У вірусів носієм генетичної інформації можуть бути як молекули ДНК, так і молекули РНК.
Розміри вірусів коливаються у межах від 8 до 750нм. Вони проходять крізь бактеріальні фільтри, а побачити їх можна тільки за допомогою електронного мікроскопа. Віруси – ультрамікроорганізми.
За формою віріонів віруси поділяються на:
паличкоподібні (віруси тютюнової мозаїки, сказу та ін.),
сферичні (віруси грипу, курячої саркоми, кору та ін.),
кубовидні (віруси натуральної віспи, папіломи людини і тварин, аденовіруси, ентеровіруси тощо),
ниткоподібні (віруси мозаїки пшениці, квасолі, сої, кавунів)
булавовидні (віруси бактерій).
Хімічний склад вірусів та їх відношення до факторів навколишнього середовища
Елементарний склад віріонів в середньому може характеризуватись такими показниками (%):
вуглець – 50 - фосфор – 0,4-0,5
кисень – 20 - сірка – 0,1-0,2
водень – 7 - зольні елементи – 2,5
азот – 16
До складу простих вірусів входить нуклеїнова кислота та білок. Складні віруси містять також ліпіди, вуглеводи та ферменти.
Нуклеїнова кислота (ДНК або РНК) становить від 1% до 40% маси віріона.
Маса білків становить 70-90%, які поділяються на структурні та не структурні.
Кількість ліпідів – 15-35%, кількість вуглеводів – 10%.
Віруси не чутливі до дії низьких температур. При дії високих температур більшість вірусів інактивується. Але вірус гепатиту термостійкий. Кислоти, луги, дезінфікуючі речовини активно знищують віруси. Ультрафіолетові промені проявляють сильну інактивуючу дію. Деякі віруси, інактивовані формаліном, зберігають імуногенні властивості, що дозволяє використовувати формалін для одержання вакцин.
Класифікація вірусів
Віруси характеризуються специфічністю, тому залежно від того, які організми вони вражають (на яких паразитують), їх поділяють на:
1) віруси рослин (фітопатогенні), які поділяються на спіральні паличкоподібні (вірус тютюнової мозаїки), спіральні ниткоподібні (х-вірус картоплі, вірус жовтухи цукрового буряка), сферичні або ікосаедричні (вірус некрозу тютюну), бацилоподібні або кулясті (вірус карликової кукурудзи, жовтої карликової картоплі, мозаїки озимиї пшениці).
Генетичним матеріалом фітопатогенних вірусів найчастіше служить РНК.
Вірусні хвороби рослин є хронічними і одужання не настає.
2) віруси, патогенні для тварин та людини. Відомо більше 500 видів. Вони містять або одноланцюгову РНК, або дволанцюгову ДНК. Деякі віруси вражають тільки один вид тварин. Потрапляють до організму людини контактним шляхом (вірус герпесу), через дихальні шляхи (віруси грипу, кору, віспи), з їжею або водою через кишково-шлунковий шлях (віруси поліомієліту, гепатиту А), через кров при укусах комах і тварин (віруси енцефаліту, сказу), при ін’єкціях (вірус гепатиту В, вірус набутого імунодефіциту)
Для лікування людей застосовують людський інтерферон.
3) віруси бактерій (бактеріофаги). Це віруси, що паразитують у бактеріальних клітинах.
Бактеріофаг – це віруси, що паразитують у бактеріальних клітинах. Більшість родин бактеріофагів містить дволанцюгову ДНК, але є такі, що мають одно ланцюгову ДНК; бактеріофаги двох родин містять одно-ланцюгову РНК. Дія фагів на бактерії строго специфічна – кожний з них викликає загибель лише певного виду бактерій. Фаги інфікують і руйнують лише молоді клітини, що розвиваються. Більшість фагів складається з багатогранної головки і хвостового відростка. Розмір головки фага 40-140 нм. Розмір хвостового відростка може досягати 250 нм в довжину і 25 нм в ширину.
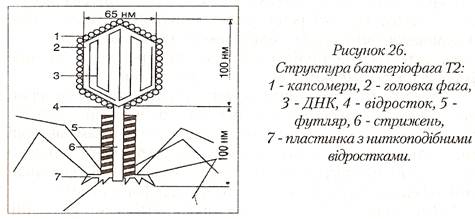
Застосування фагів. Основане на їх суворій специфічності і властивості руйнувати мікробні клітини або вступати з ними у симбіоз. В наш час фаги широко застосовують при лікуванні і профілактиці стафілококових і стрептококових захворювань, особливо таких, які не піддаються дії антибіотиків, а також холери і чуми. Фагопрофілактику проводять у період підйому захворюваності і загрозі виникнення епідемій. Немає протипоказань до застосування фага, вони не шкідливі для людей і діють вибірково, лізуючи лише певні мікроби. Проводиться індивідуальний підбор фага. Фаги випускаються фармакологічною промисловістю у рідкій та сухій формі. Фаги використовують для ідентифікації виду бактерій – збудників інфекційних захворювань.
У промисловості фаги можуть вражати клітини бактерій та інших мікроорганізмів. Бактеріофаги приносять велику шкоду молочній промисловості.
Основні шляхи попередження розвитку бактеріофага: підтримка асептичних умов вирощування заквасок, заміна заквасок, чергування в заквасках штамів, які не чутливі до великої кількості типів бактеріофагів та ін.
Практичне застосування фагів. Застосування фагів базується на їх строгій специфічності і здатності руйнувати мікробні клітини або вступати з ними в симбіоз.
1. Фагопрофілактика та фаготерапія. Попередження та лікування інфекцій за допомогою фагів базується на тому, що зустрівши в організмі хворого збудника захворювання, фаг знищує його.
В даний момент фаги широко застосовують при лікуванні та профілактиці стафілококових та стрептококкових захворювань, особливо таких, які не піддаються дії антибіотиків. Цінним фактором являється те, що нема протипоказань до застосування фагів, вони нешкідливі для людей та діють вибірково, тобто лізують тільки визначенні мікроорганізми.
Однак застосування фагів для лікування захворбвань незавжди ефективно. Це обумовлюється великою кількістб фаготипів одного й того ж збудника. Що потребує індивідуального добіру фагу.
2. Фагодіагностика та ідентифікування видів. Використовують відомі (діагностичні) фаги для ідентифікації виа культур бактерій – збудників інфекціонних захворювань. Індивідуальні культури відповідають тому фагу, який її лізував. Сувора специфічність дає можливість типувати варіанти усередині виду.
Фізіологія мікроорганізмів
Вивчає життєві функції мікроорганізмів:
обмін речовин,
живлення,
дихання,
ріст і розвиток,
розмноження,
реакції на зовнішні подразники,
пластичність,
інтенсивність взаємодії з факторами середовища.
Фізіологія являється науковою основою промислового використання мікроорганізмів у мікробіологічних виробництвах біологічно активних речовин (БАР), ферментів, вітамінів, антибіотиків, амінокислот, органічних кислот.
Мікроорганізми здатні до синтезу різних біологічно активних речовин, а деякі з них – до надмірного синтезу окремих речовин – метаболітів, ці мікроорганізми звуть продуцентами. Чиста культура – продуцент певної БАР є головним компонентом мікробного синтезу. Показниками культури, її якості є ступінь і термін накопичення цільової речовини, висока швидкість росту, чутливість до джерел вуглецю і супутних сполук живильного середовища. Мікробіологічне виробництво – багатостадійний процес підготовки, ферментації, сепарування, вилучення цільового продукту в асептичних умовах.
Хімічний склад мікроорганізмів
Хімічний склад клітин мікроорганізмів практично не відрізняється від складу клітин інших живих істот біосфери. Основну частину маси вегетативної клітини складає вода – 70-90%, вміст сухої речовини від 10 до 30%. Суха частка мікробної клітини на 85-97% представлена органічними сполуками, на 3-15% - мінеральні речовини.
Вода буває в двох станах: вільна і зв’язана. Вільна – розчинник, джерело іонів Н+, ОН-, учасник ферментолізу, транспорту речовин. Друга– існує в 4-ох видах – хімічно-, адсорбційно-, осмотично- і механічно зв’язана вода.
Вміст води в клітинах мікроорганізмів впливає на їх життєздатність, їх стан, тому втрата вільної води переводить клітину до анабіозу, а зв’язаної – до загибелі клітин (абіозу).
Вміст білків складає 50-80% сухої речовини, вони є основними і нийважливішими біополімерами в біомасі мікроорганізмів. Білки являються будівельним матеріалом і виконують катлітичну, транспортну, регулюючу, антигенну функції. Білки – ферменти забезпечують протікання біохімічних реакцій. В залежності від їх локалізації розрізняють екзоферменти, які клітина продукує назовні для забезпечення позаклітинного травлення, та еноферменти – каталізатори внутрішньоклітинного метаболізма. Набор ферментів являється характеристикой певних видів. Здатність синтезувати конститутивні ферменти кодується клітинним геномом, являється постійною ознакою, і не залежить від субстрату. Адаптивні або індуцибельні ферменти синтезуються при зміні факторів середовища. Нуклеопротеїди і нуклеїнові кислоти відповідають за процеси спадковості і мінливості.
Прості і складні вуглеводи виконують запасну, енергетичну. Захисну, будівельну, антигенну та інші функції.
Ліпіди мікроорганізмів входять до складу біомембран і клітинних стінок, являються джерелом енергії, виконують функцію запаса. Від концентрації ПНЖК залежить резистентність деяких бактерій до дії високих і низьких температур.
Здатність окремих видів продукувати пігменти використовується для ідентифікації видв мікроорганізмів. Існують бактерії, які не здатні синтезувати вітаміни, але їх життєдіяльність залежить від концентрації факторів росту в середовище. Такі мікроорганізми називають ауксотрофами. Певні види молочнокислих бактерій застосовують як живі тест – культури для визначення концентрації вітамінів.
Кількісно, в залежності від критерія, елементи поділяють на макро-, мікро- і ультрамікроелементи. Основу органічних речовин складають біоелементи або органогени. До органічних сполук мікроорганізмів відносяться: білки, нуклеїнові кислоти, вуглеводи, ліпіди, вітаміни, токсини, пігменти. Вміст білків складає 50-80% сухої речовини, вони є будівельним матеріалом клітини. Вміст нуклеїнових кислот у клітині залежно від виду бактерій і поживного середовища коливається від 10 до 30 % на суху речовину. Концентрація вуглеводів становить 12-28%, вони використовуються клітинами для синтезу різних сполук та як джерело енергії. Ліпіди складають 3-10% на суху речовину, і виконують функції енергетичного матеріала або запасних поживних речовин.
Види обміну речовин у мікроорганізмів
Важлива, суттєва, віждміна риса мікробіоти – метаболізм - сукупність процесів, що відбуваються у клітинах в процесі життєдіяльності. Він складається з двох різноспрямованих потоків реакцій: конструктивного та енергетичного метаболізму.
Конструктивний обмін (анаболізм) має місце, коли з речовин, що надходять зовні, та енергії синтезуються речовини клітини.
Енергетичний обмін (катаболізм ) – реакції розкладання органічних субстратів з утворенням макроергічних зв’язків АТФ. Енергетичнй та конструктивний метаболізми одночасні, тісно пов’язані. Виділення енергії відбувається в процесі дихання під впливом окисно-відновних ферментів.
Усі бактерії за типом дихання діляться на аеробів та анаеробів.
Аероби - бактеріальні клітини, які окислюють органічні субстрати з виділенням енергії за реакцією: С6Н12О6+6О2=>6СО2+6Н2О+Q. Енергія, яка вивільняється, застосовується бактеріями на біосинтез, структур клітини, механічну роботу або виділяється у вигляді світової та теплової енергії. При термогенезі бактеріальні клітини продукують назовні теплову енергію. Це явище виникає при самозігріванні або самозапалюванні зерна з низькою теплопровідністю при t=80-90 0С. Мікроорганізми які здатні перетворювати хімічну енергію макроергічних зв’язків АТФ в світло називаються фотогенними. (Photobacterium phosphoreum)
Анаероби – бактерії, які не можуть застосовувати атмосферний кисень. Для одержання енергії вони використовують анаеробні дегідрогенази. Цей тип відкрив Пастер і дав назву бродіння (спиртове, маслянокисле).
Розрізняють підтипи: облігатні; факультативні типи дахання. До Облігатні аеробів відносяться сарцини, вібріон холери, micromycetes, плівчасті дріжджі, mycobacterium. Факультативних аеробів не виявлено. Мікроаерофіли потребують кисень в незначній кількості (Thamnidium, Cladosporium). Облігатних анаеробів в лабораторії культивують на середовищі Китт-Тароцці. До них відносяться: , Clostridium sporogenes, Clostridium putrificum, Escherichia coli i Proteus vulgaris. Вони являються факультативними анаеробами.
Типи живлення
У класифікації типів живлення використовується сучасна термінологія, запропонована у 1946 році на мікробіологічному симпозіумі. На базі трьох критеріїв розглядається кілька типів живлення:
щодо джерела вуглецю (автотрофія, гетеротрофія);
щодо донора електронів (органотрофія, гетеротрофія);
щодо джерела енергії (хемотрофія, фототрофія).
За відношенням до джерела вуглецю мікроорганізми поділяються на автотрофи та гетеротрофи. Автотрофи застосовують СО2 і самі синтезують органічні речовини. Донором електронів в них є неорганічні сполуки: Н2, NH3, H2S, S, Fe2+. В залежності від донора бактерії мають назву : водневі, нітрифікуючи, сіркобактерії, залізобактерії. Їх назва – літоавтотрофи. Більшість з них виконує важливі функції в кругообігу речовин в природі. С.М. Виноградський відкрив новий тип живлення – хемосинтез,коли окислення молекулярного водню, NH3,H2S і задіза є джерелом енргії аеробних актерій.
Серед автотрофів відомі ціанові, пурпурові, зелені актерії, я кі використовують сонячну енергію і проводять фотосинтез.
Гетеротрофи – це мікроорганізми, які для живлення потребують органічні сполуки: амінокислоти, білки, вуглеводи, ліпіди. Їх сучасна назва – хемоорганогетеротрофи. У цих організмів вуглецева сполука є єдиним джерелом вуглецю, електронів і енергії. Це найбільш досліджена група мікроорганізмів, широко розповсюджена в природі. Вони поділяються на сапрофіти і паразити.
Враховуючи три критерії виділяють 8 груп мікроорганізмів, що різняться за типом живлення
| Джерело енергії | Джерело електронів | Джерело вуглецю | Група мікроорганізмів |
| Світло | Неорганічні сполуки | СО2 | Фотолітоавтотрофи |
| Світло | Неорганічні сполуки | Органічні сполуки | Фотолітогетеротрофи |
| світло | Органічні сполуки | СО2 | Фотоорганоавтотрофи |
| Світло | Органічні сполуки | Органічні сполуки | фотоорганогетеротрофи |
| Хімічні реакції | Неорганічні сполуки | СО2 | Хемолітоавтотрофи |
| Хімічні реакції | Неорганічні сполуки | Органічні сполуки | Хемолітогетеротрофи |
| Хімічні реакції | Органічні сполуки | СО2 | Хеморганоавтотрофи |
| Хімічні реакції | Органічні сполуки | Органічні сполуки | Хемоорганогетеротрофи |
Механізм транспорту поживних речовин до клітини
Мікроорганізмам властивий голофітний тип живлення. Вони не мають органів травлення, а поживні речовини надходять у водному розчині різними механізмами. Особлива регуляторна роль в транспорті поживних речовин до середини клітини і виведенні метаболітів назовні належить цитоплазматичній мембрані. ЦПМ має пори малого діаметру, які характеризуються вибірковою напівпроникністю. В ЦПМ локалізовані пермеази – білки переносу із суворою специфічністю до субстратів. Їх кількість буває значною.
Розрізняють такі типи транспорту:
активний транспорт – коли з середовища із низькою концентрацією поживних речовин у клітину поступають харчові речовини за допомогою пермеаз і енергії АТФ.
полегшена дифузія – коли із середовища з високою концентрацією речовини транспортуються за допомогою пермеаз без затрат енергії.
пасивна дифузія – коли із середовища речовини рухаються з зони високої концентрації в зону низької концентрації в клітину за градієнтом концентрації або електростатичного потенціалу.
Існують три типи станів клітини, які обумовлені співвідношенням концентрації речовин назовні і в середині клітини.
1-ий стан – нормальний стан клітини – тургор – коли клітина знаходиться в ізотонічному розчині (0,85% NaCl). В цьому випадку дифузія осмотично активних речовин в клітину формує внутрішньоклітинний осмотичний тиск. Він зветься тургор і коливається у великих діапазонах 1,5 МПа. Тургор забезпечує транспорт води у клітину. ЦПМ щильно прилягає до клітинної стінки і тиск в середині клітини трохи більший ніж тиск ззовні.
2-ий стан – плазмоліз – якщо клітина потрапляє в середовище з високим осмотичним тиском (гіпертонічний розчин), то ЦПМ віддає воду і вміст клітини зневоднюється, стискається. Клітина переходить спочатку у стан анабіозу, тобто зберігає життєздатність, а потім – до стану абіозу, гине. В технології консервування застосовують при солінні 25% NaCl і при виготовленні варіння 50% сахарозу. В такій концентрації вони виявляють бактеріостатичну дію, тобто бактерії не розмножуються але не гинуть і бактерицидну дію, коли наступає загибель бактерій.
3-ий стан – плазмоптіс – бактеріальна клітина потрапляє в гіпотонічний розчин, наприклад, це відбувається коли клітина потрапляє в дистильовану воду.
Існують мікроорганізми, які виявляють резистентність до високої концентрації речовин в середовищі. Стійкі організми, що можуть жити, розвиватись, розмножуватися при високому осмотичному тиску називаються осмофільними. Відомі бактерії галофіли, які здатні розмножуватися в середовищі з високим вмістом NaCl, серед них бувають облігатні галофіли, які потребують обов’язково 12% NaCl і більше. Факультативні галофіли можуть розмножуватися і при 1-2%,і 10-12%.
Розмноження мікроорганізмів
Ріст бактерій – це збільшення кількості компонентів в клітині, її розмірів і маси. Розмноження бактерій – це збільшення числа клітин в популяції. Прокаріоти розмножуються бінарним діленням. Також відомі факти кон’югації ( статтєве розмноження). В штучних умовах на незмінному поживному середовищі розмноження відбувається за певними закономірностями.
Розрізняють 4 фази розмноження:
лаг-фаза або фаза затримки росту. Вона триває дві години. Це відбувається тому що бактерії пристосовуються до умов, кількість клітин впродовж цієї фази не збільшується.
фаза логарифмічного росту – при цьому в геометричній прогресії збільшується кількість клітин. Вона ще називається експоненціальною і триває 5-6 годин.
стаціонарна фаза – це коли кількість нових клітин дорівнює кількості відмерлих клітин. Ця фаза триває 2 години.
фаза відмирання – в цій фазі кількість нових клітин менше ніж кількість відмерлих. У спороутворюючих мікроорганізмів йде процес утворення спор.
До причин загибелі мікроорганізмів відносяться лізіс власними ферментами ( цей процес проходить у лізосомах), накопичення токсинів-метаболітів життєдіяльності та збіднення поживного середовища.
Біогеохімічна діяльність мікроорганізмів. Роль мікроорганізмів у кругообігу речовин у природі
Вміст в атмосфері азоту, кисню і вуглекислого газу, інших хімічних елементів, виявлених на поверхні Землі і необхідних для життя, відображає рівновагу між їх утворенням у біологічних і геологічних процесах. Ці перетворення відбуваються у всій біосфері, тобто в тій тонкій оболонці життя на поверхні Землі, що охоплює океани, моря, прісні водоймища, грунт континентів і нижню частину атмосфери, і в якій тільки і містяться живі організми. Загального вмісту головних хімічних елементів, необхідних для життя, зокрема вуглецю й азоту, які є в атмосфері, при їхньому однобічному споживанні навряд чи вистачило б на мільйони років.
Біосфера знаходиться в більш-менш стійкому стані завдяки безперервному припливу сонячної енергії і постійному кругообігу вуглецю кисню, азоту, сірки і фосфору. У цілому ці процеси виглядають так: за допомогою сонячної енергії фотосинтезуючі організми перетворюють вуглекислий газ та інші неорганічні речовини в глюкозу та інші органічні сполуки, які прямо чи побічно служать джерелом енергії для всіх інших організмів. У свою чергу фотосинтезуючі організми – одноклітинні водорості, які живуть в океані і вищі рослини, які ростуть на суші, - служать джерелом харчування для тварин. У такий спосіб органічна речовина, що накопичується щорічно, у процесі фото синтезуючої діяльності переробляється на різних рівнях життя консументами і деструкторами. До перших належать, головним чином, тварини, до других – гриби і бактерії. Послідовність цих подій виражається в трофічних ланцюгах або ланцюгах харчування. Кінцева, деструктивна ланка цього ланцюга – мінералізація органічних речовин з поверненням вуглекислого газу в атмосферу – здійснюється гетеротрофними мікроорганізмами.
Деструктаторами природних біополі мерів (білки, нуклеїнові кислоти, геміцелюлози, пектин, крохмаль, целюлоза та ін.) можуть бути лише ті мікроорганізми, які синтезують гідролітичні ферменти. В аеробній зоні до таких мікроорганізмів відносяться гриби, грампозитивні бактерії, у тому числі й актиноміцети. В анаеробній зоні – це тільки бактерії, в основному з групи клостридій. В аеробній зоні відбувається практично повне перетворення полімерів із звільненням вуглекислого газу. В анаеробних умовах у процесі первинного розкладання органічних речовин як продукти розпаду утворюються жирні кислоти, спирти і молекулярний водень, які частково використовуються сульфітредукуючими, денітрифікуючими і метаноутворюючими бактеріями. Володіючи досить активним ферментним апаратом, мікроорганізми здійснюють процеси розщеплення і синтезу самих складних органічних речовин. Завдяки їх мінералізуючій діяльності поверхня Землі постійно очищується від трупів тварин і залишків рослин. За словами В.Л. Омелянського, мікроорганізми є дійсними могильниками органічного світу.
Кругообіг азоту
Азот – це біогенний елемент, складова частина структурних білків, ферментів, нуклеїнових кислот, АТФ та ін. Близько 4/5 повітря, яке нас оточує, приходиться на частку вільного азоту, над кожним гектаром ґрунту піднімається стовп повітря, котрий містить близько 80 тис. тонн молекулярного азоту. Газоподібний вільний азот не асимілюється рослинами, тваринами і людиною. Багато форм зв’язаного азоту не можуть служити джерелом азотного живлення для рослин. Азот, який надходить у виді білкових речовин у грунт разом із залишками рослин і тварин, зовсім не підходить для цих цілей, він має бути окислений у солі азотистої та азотної кислот (нітрити і нітрати). Кругообіг азоту – це складний біохімічний процес, який складається з чотирьох етапів: азот-фіксація; амоніфікація; нітрифікація; денітрифікація.
Фіксація атмосферного азоту здійснюють мікроорганізми, які здатні засвоювати молекулярний азот N2 і переводити його в зв’язаний стан. Внаслідок їх діяльності йде накопичення рослинних білків, в цьому приймають участь два типи бактерій:
бульбачкові – вони живуть у симбіозі з бобовими рослинами;
другі живуть вільно у грунті і воді. (Azotobacter, Clostridium pasturiaum)
Рослинні білки поглинають споживачі органічних речовин консументи. Результат – накопичення тваринних білків.
Амоніфікація – це розкладання редуцентами білків рослинного походження як мертвих, так і живих. Сапрофіти проводять амоніфікацію мертвих органічних сполук. Це бактерії і мікроміцетові гриби. Назва процесу походить від аміак NH3, тому що в процесі амоніфікації утворюється аміак як кінцевий продукт гідролізу, мінералізації білків в аеробних і анаеробних умовах під дією протеолітичних ферментів гнильних бактерій. У процесі гниття йде очищення Землі від трупів тварин і залишків рослин. Гнильні бактерії виконують санітарну роль. В аеробних умовах кінцеві продукти гниття – CO2, H2O, NH3, H2S, мінеральні солі. В анаеробних умовах поруч з CO2, H2O, NH3, H2S утворюються скатол, індол, крезол, меркаптани, леткі жирні кислоти, діаміни – тобто отруйні речовини. Серед гнильних бактерій є патогенні (Bac.cereus) і умовно патогенні Escherichia coli, Proteus vulgaris.
Гнильні бактерії – це організми, які мають протеолітичну активність. Вони за своєю природою є амоніфікаторами, які мають оптимум температурного розвитку в межах 30 – 40 ºС. Тобто їх відносять до мезофілів. Вони є хемоорганогетеротрофами і за морфологією є паличкоподібними, але серед них є палички, які утворюють спори, не утворюють спор, аеробні та анаеробні. Тому виділяють 4 группи:
Аеробні палички, що не утворюють спор. Представники групи - грамнегативні та рухливі палички - Pseudomonas fluorescens i Serratia marcescens. Ці два вида утворюють пігменти: перший флюорисцентний, а другий – пігмент червоного кольору . На МПЖ проводять гідроліз желатину, згортають молоко,утворюють аміак, сірководень
Аеробні палички, що утворюють спори. Вони грампозитивні та рухливі. До них належать: Bac. subtilis, Bacillus licheniformis, Bac. cereus, Bac. megaterium, Bac. mycoiges. Ці бактерії здатні проводити гідроліз желатину, згортають молоко.
Факультативні анаероби. Бактерії нездатні утворювати спори, грамнегативні, рухливі. Представники: Escherichia coli, Proteus vulgaris. Протеї на скошеному агрі утворює вуаля – фенмен роїння. Кишкова паличка не являється типовим гнильним мікроорганізмом.
Облігатні анаероби. Вони здатні утворювати спори, грампозитивні, рухливі.
Предстаники: Clostridium sporogenes i Cl.рutrificum.
Ці мікроорганізми здатні накопичувати аміак і гідролізують МПЖ , але при цьому вони утворюють велику кількість інших газів (CO2, H2S). Вони можуть складати залишкову мікрофлору в стерилізованих консервах, і викликати псування, яке називається бомбаж. Всі гнильні бактерії виконують роль санітарів. Вони являють собою фактори біологічного самоочищення грунту, води. Їх застосовують в работі споруджень по знешкодженню фікальних нечистот і стічних вод, полів асенізації і полів зрошення.
Слідуючим етапом кругообігу азоту є нітрифікація. Це перетворення аміаку, амонійних солей до нітритів і нітратів. За схемою NH3→NO2 →NO3.На першому етапі приймає участь Nitrosomonas, а на другому Nitrosospira. Ці бактерії відносяться до хемолітоавтотрофів, вони проводять хемосинтез, є анаеробами, їх зуть нітрифікуючими. При окислені амаіка до азотистої і азотної кислот вивльняється енергія.
Денітрифікація – це процес протилежний нітрифікації:
HNO3 2HNO2 + O2
2HNO2 - проміжне з’єднання + O2
Проміжкове з’єднання N2+ H2O +O2
Цей процес відбувається під дією мікроорганізмів Pseudomonas denitrificans. Ці процеси призводять до збіднення грунту мінеральними солями, але їх діяльність компенсується діяльністю азот фіксуючих мікроорганізмів.
Кругообіг вуглецю
Неорганічний вуглець надходить в органічну форму біосфери в процесі фотосинтезу і навпаки – органічний вуглець повертається в атмосферу у вигляді СО2 завдяки різноманітним перетворенням природи. Велика роль в цьому належить діяльності мікроорганізмів при бродінні. Кожний тип бродіння спричиняється певною групою мікроорганізмів, які використовують вуглеводи, то їх називають хемоорганогетеротрофи. Збудники бродінь широко розповсюджені в природі і мають народногосподарське значення. Крім того, запаси вуглецю в атмосфері поповнюється за рахунок вулканічної діяльності і спалювання людиною паливних копалин. Хоча основна частина діоксиду вуглецю потрапляє в атмосферу поглинається океаном і відкладається у виді карбонатів, вміст діоксиду вуглецю у повітрі повільно, але неухильно підвищується.
При спиртовому бродінні відбувається розщеплення глюкози ферментами дріжджів, деяких грибів ( р. Mucor) з утворенням етанолу в анаеробних умовах. Спиртове бродіння застосовується при виробництві спирту, гліцерину, вина, пива, квасу, хліба, квашених овочів і ін. сферах.
Молочнокисле бродіння – це процес розпаду глюкози до молочної кислоти під дією ферментів молочнокислих бактерій. За рядом характеристик – за морфологією та культуральними ознаками та ін. їх класифікують на групи, але їх всіх об’єднує біохімічні властивості – здатність проводити бродіння – сахаролітична активність.
В залежності від того, які утворюються метаболіти, бродіння розрізняють: гомоферментативне, гетероферментативне, біфідобродіння
Але для всіх груп є інші загальні характеристики - вони всі відносяться до хемоорганогетеротрофами, ауксотрофні і мікроаерофіли. При гомоферментативному бродінні утворюється один метаболіт – молочна кислота.
При гетероферментативному бродінні утворюється декілька метаболітів: молочна кислота, етанол, оцтова кислота, СО2 і сукцинат. При біфідобродіння утворюється два метаболіти – молочна і оцтова кислоти.За морфологією молочнокислі – це гетерогенна група. Серед них зустрічаються палички і коки, але всі вони неспороутворюючи, грампозитивні і нерухливі.
Представники:
До гомоферментативних бактерій відносяться: Streptococcus lactis – мезофіл,Streptococcus casei – мезофіл; а до термофілів відносяться - Lactobacillus bulgaricus – термофіл, Lactobacillus acidophilus – термофіл
До гетероферментативн бактерій відносяться: Leuconostoc mesenteries, Lactobacillus fermentum
До біфідобактерій відносяться: Bifidobacterium bifidum
Збудники молочнокислого бродіння зустрічаються в ґрунті, на рослинах, в молочнокислих продуктах, в складі біоценозу кишкового тракту. До 50 % маси фекалій – це молочнокислі та інші бактерії. Вони являються антагоністами гнильних, тому що вони синтезують антибіотичні речовини – нізин, бревін, які є неблагоприємними до розвитку гнильних. накопичують молочну кислоту.
Молочнокислі бактерії використовують органічні форми азоту як джерело азотного харчування. Багато молочнокислих бактерій можуть асимілювати білки, хоча краще розвиваються на амінокислотах, пептидах і полі пептидах. Продукти розподу білкової молекули прекрасно засвоюються цими бактеріями.
Маслянокисле бродіння відбувається в анаеробних умовах. Під дією ферментів бактерій відбувається процес перетворення вугеводів переважно до масляної кислоти, води та вуглекислоти.Поруч утворюються бутанол, ацетон, етанол, оцтова кислота. Збудники маслянокислого бродіння відносяться до роду Cl. Це грампозитивні палички, рухливі, спороутворюючи (форма клітин ракетки, веретено). Клострідії – облігатні анаероби, хемоорганогетеротрофи, мезофіли, чутливі до рН середовище (opt. Для них рН = 7,0-7,3)
Як джерело вуглецю маслянокислі бактерії можуть використовувати моно- і дисахариди, деякі полісахариди (декстрини, крохмаль), молочну і піровіноградну кислоти, маніт, гліцерин і інші сполуки. У складних білкових середовищах при відсутності вуглеводу маслянокислі бактерії погано ростуть або зовсім не ростуть. Джерелом азоту служать різноманітні речовини – амінокислоти, аміачні сполуки і навіть молекулярний азот. Серед маслянокислих бактерії є сапрофіти (C. pasteurianum, C.buturicym, C. felsineum) та патогенні види (C. botulinum, C. tetani, C. perfringens).
Маслянокисле бродіння іноді буває небажаним. Наприклад, при його розвитку в кормах, що заквашуються, білкова частина корму розкладається, а масляна кислота, що накопичується, додає йому неприємного запаху. Разом з тим масляна кислота потрібна для деяких промислових цілей. Її одержують на заводах, піддаючи бродінню спеціально підготовлені середовища чистою культурою маслянокислих бактерій. Кислоту, що утворилася, відокремлюють і очищають хімічним методом.
Вплив факторів зовнішнього середовища на життєдіяльність мікроорганізмів
Успішна боротьба з мікроорганізмами й ефективне використання їх у виробничих процесах можливі за умов розуміння їх взаємодії із зовнішнім середовищем. Застосовуючи мікроорганізми для виробництва харчових продуктів, недостатньо тільки вибрати активні культури, необхідно створити оптимальні умови для їхнього розвитку. Регулюючи умови існування, можна не тільки керувати життєдіяльністю мікроорганізмів, але й викликати бажані їх зміни.
Фактори поділяються на три групи:
Фізичні
Хімічні
Біологічні
Фізичні фактори: температура, вологість, осмотичний тиск або концентрація розчинених речовин у середовищі, радіоактивні випромювання, радіохвилі, магнітне поле, гідростатичний тиск, вплив невагомості, промениста енергія, ультразвук.
Температура
Механізм регуляції температури клітин у мікроорганізмів відсутній, тому що для різних груп виявлено температурний діапазон. Він характеризується трьома кардинальними точками : мінімум, оптимум, максимум.
Мінімальна температура – це така, нижче якої припиняється ріст і розмноження бактерій. Вони переходять спочатку в анабіоз і абіоз.
Оптимальна температура відповідає найвищий швидкості росту і розмноження мікроорганізмів.
Максимальна температура – це та, вище якої ріст неможливий, бактерії переходять в стан анабіозу, а потім – абіозу.
В залежності від трьох кардинальних точок бактерії розділяють на групи:
| min | opt | max | |
| психрофіли | -7 | 10 | 20 |
| мезофіли | 10 | 30 | 45 |
| термофіли | 40 | 60 | 75 |
Представники психрофілів: Thamnidium, Penicillum, Pseudomonas. Живуть в холодних морях і на крайній півночі.
Представники мезофілів: Escherichia coli, C. Botulinum, Bac. Subtilis. Всі патогенні, гнилостні.
Представники термофілів: Lactobacillus bulgaricus та ін. Зустрічаються в гарячих джерелах, в торфі, в грунті.
На згубній дії високих температур засновано багато прийомів знищення мікроорганізмів у харчових продуктах і в різних інших об’єктах, наприклад, кип’ятіння, варіння, обсмажування, бланшування продуктів харчування, стерилізація, обробка паром виробничого устаткування.
У харчовій промисловості широко застосовують два способи впливу високих температур на мікроорганізми: пастеризацію і стерилізацію.
Пастеризація – це нагрівання продукту до 90 – 100 ºС, тривалість процесу залежить від температури і звичайно складає 20 – 40 хв. При пастеризації гинуть не всі мікроорганізми. Деякі термостійкі бактерії, а також спори багатьох бактерій залишаються живими. У зв’язку з цим пастеризовані продукти варто охолоджувати до температури не вище 10 ºС і зберігати на холоді, щоб затримати проростання спор і розвиток збереження клітин. Пастеризують молоко, пиво, ікру, фруктові соки і деякі інші продукти.
Стерилізація – це нагрівання при температурах вище 100 ºС протягом певного часу, при якому відбувається загибель вегетативних клітин мікроорганізмів і їхніх спор. Стерилізації піддають різні банкові консерви, багато предметів і матеріалів, які використовуються в медичній і мікробіологічній практиці.
Вплив холоду
Холодильна технологія базується на постулаті, що більшість мікроорганізмів (мезофіли і термофіли) не здатні розмножуватися при температурі нижче нуля. В промисловості застосовуються два способи зберігання під дією холода – в охолодженому стані не вище 4 ºС і при заморожуванні не вище -8 ºС. Але відомі факти, що деякі мезофільні організми витримують тимчасово низькі температури. Наприклад, Clostridium Botulinum зберігається протягом одного року при температурі – 16 ºС. Існують спори, які витримують температуру – 190 ºС, а в природних кригах знайдено життєздатні мікроорганізми, які перебували там 12 тисяч років.
Фактори загибелі мікроорганізмів під дією холоду
Механічна дія кристалів льоду.
При малій швидкості тепло відводу і повільному тривалому заморожуванні часом 10 хвилин до температури -80 ºС в міжклітинному просторі утворюються великі кристали. При цьому гине частина мікроорганізмів. При швидкому заморожуванні за короткий термін ( до 20 хвилин) і у середині клітини і між ними утворюються дрібні кристали, які разом формують суцільні моноліти льоду. Більшість мікроорганізмів гине. В даному випадку це явище зветься вітрифікацією.
Осмотичний тиск.
Внаслідок виморожування води, солі, монози концентруються і в середовищі утворюється гіпертонічний розчин, підвищений осмотичний тиск в оточуючим середовищі який спричиняє плазмоліз бактерій і вони переходять у стан анабіозу і потім абіозу.
Вологість.
Зменшення показника „ водна активність” і доступної води гальмує фізіологічні процеси – розвиток, метаболізм, дихання трофіїв. Бактерії переходять в анабіоз і гинуть.
Однак заморожування не спричиняє стерилізуючої дії. У заморожених продуктах завжди є живі життєздатні мікроорганізми. Під час розморожування продуктів, особливо при витіканні з них соку, мікроорганізми знову розмножуються і викликають псування. Тому відтаювати заморожені харчові продукти слід безпосередньо перед вживанням.
Холодостійкість
Холодостійкість – це відношення мікроорганізмів до впливу низьких температур. Психрофільні бактерії мають мінімальну кардинальну точку – 12 ºС – нижче неї вони не розмножуються і переходять у стан анабіозу. Механізм адаптації до стану холоду у різних мікроорганізмів різноманітні. У деяких Грамнегативних в цитоплазматичній мембрані і в стінці клітини є велика кількість ліпідів. Стабілізація мембран відбувається за рахунок підвищення в них концентрації полі ненасичених жирних кислот, а резистентність спор Грампозитивних до дії холода пов’язана з низькою концентрацією вологи.
Термостійкість
Термостійкість – відношення мікроорганізмів до температури, які перевищують максимальну кардинальну точку. Термостійкість спор пов’язана з малою концентрацією води та наявністю дипіколінату Са. У деяких термостійкість пов’язана з низькою концентрацією полі ненасичених жирних кислот.
В харчовій промисловості застосовують два методи – пастеризація і стерилізація.
Ефективність стерилізації залежить від початкового вмісту бактерій: чим кількість їх більше, тим більше залишається живих клітин у продукті і вони викликають псування.
Вологість
У вегетативних клітинах бактерій міститься до 85% води, з нею транспортуються поживні речовини у середину, а метаболіти – назовні . Мікроорганізми розвиваються тільки в субстратах з вільною водою, а зв’язані форми води для них недоступні. При видаленні вологи нижче мінімального рівня розмноження припиняється. В залежності від критерію мінімальної потреби мікроорганізмів в волозі і доступність води мікроорганізми класифікують на групи:
гідрофіти
мезофіти
ксерофіти
| Групи мікроорганізмів | Представники | Активність води | Відносна вологість |
| Гідрофіти | бактерії | 0,94-0,90 | 95-90 |
| Мезофіти | Гриби, дріжджі | 0,88-0,8 | 90-80 |
| Ксерофіти | Aspergillus, Micrococcus | 0,75-0,65 | 75-65 |
Водна активність харчових продуктів знаходиться в тих же межах, що і у більшості бактерій, тому вони утворюють сприятливі умови для розвитку бактерій, тому продукти псуються. Продукти, у яких активність води менша ніж 0,7 можуть довгостроково зберігатися без мікробного псування. В технології застосовується зберігання продуктів харчування у вигляді сухих продуктів. Але вони не є стерильними і в них є залишкова мікрофлора: дріжджі, молочнокислі і коки.
Здавна застосовується зберігання різних харчових продуктів у сухому вигляді. Сухі продукти не є стерильними і завжди містять різні мікроорганізми, серед яких можуть бути і патогенні форми. У висушеному стані багато мікроорганізмів зберігаються життєздатними протягом тривалого часу. Наприклад, червоно тифозні бактерії, більшість стафілококів і мікрококів, молочнокислі бактерії можуть зберігатися в сухому вигляді тижнями і місяцями, а оцтовокислі бактерії відмирають швидко. Стійкі до висушування дріжджі. Спори бактерій у висушеному стані зберігають здатність до проростання протягом десятків років. Зволоження сухих продуктів у період зберігання може спричинити їх псування внаслідок розвитку збережених на них мікроорганізмів.
Сублімаційне сушіння продуктів (висушування у вакуумі із замороженого стану) сприяє значно кращому збереженню їх харчової якості. Однак мікроорганізми добре переносять таке висушування і навіть після багаторічного перебування в цьому стані зберігають життєздатність. Тому до продуктів, що піддаються такому висушуванню, варто пред’являти суворі санітарно-гігієнічні вимоги.
Концентрація розчинних речовин у середовищі або осмотичний тиск
Внутрішньоклітинний осмотичний тиск у різних мікроорганізмів коливається в широких межах. В одних, наприклад, у багатьох збудників псування харчових продуктів, він складає 0,5 – 1,5 МПа, в інших – значно більше. Величина осмотичного тиску клітинного соку деяких плісеневих грибів з роду Aspergillus досягає 20 МПа, ґрунтових бактерій 5 – 8 МПа. Навіть у представників одного виду, що живуть у різних умовах, внутрішньоклітинний осмотичний тиск може значно коливатися.
В пріроді мікроорганізми зустрічаються в субстратах з різноманітним вмістом розчинених речовин, а, отже, з різним осмотичним тиском. Мікроорганізми, що можуть жити при високому осмотичному тиску, називаються осмофільними.
Бактерії-галофіли можуть рости на середовищах з високим вмістом солей. Облігатні галофіти обов’язково потребують високої концентрації NaCl, факультативні галофіти нормально розмножуються при різних концентраціях хлориду натрію. Галофітами являються мікрококи, лактобацили, деякі микроміцети, вібріони, спірили.
В нормі внутрішньоклітинний тиск бактерій, який зветься тургор, перевищує осмотичний тиск середовища і досягає 1,5 МПа або 15 атмосфер.
Існує три стану бактерій в залежності від осмотичного тиску:
плазмоліз
плазмоптис
тургор
А також бактерій на осмофільних і галофільних
Промениста енергія
Промениста енергія – це енергія у вигляді електромагнітних хвиль. Різні спектри променистої енергії мають різну довжину хвиль, яка визначає їх характер і властивості.
Електромагнітні хвилі поділяються на групи:
радіохвилі – у них найбільша довжина хвилі 1500 нм;
світлові промені поділяються на три підгрупи: інфрачервоні (760 нм), видимі (760-380 нм), ультрафіолетові промені (380-200 нм);
іонізуюча радіація – найбільш коротка хвильова частина спектру.
Дія на прокаріоти різних видів випромінювання залежить від їх енергії і дози випромінювання. Ультрафіолетові випромінювання використовується для дезінфекції повітря в холодильних камерах, виробничих приміщеннях. За 6 годин ультрафіолетове опромінення знищують 80% бактерій і грибів. Використовують ультрафіолетове опромінення при розливі, фасуванні, пакуванні продуктів, для обробки питної води, знезаражування тари. Можливе застосування для стерилізації соків в тонкому шарі. До ультрафіолетових променей стійкі коки, які містять каротиноїди, деякі види грибів та бактерій. В харчовій промисловості застосовують гамма-промені, промені високої та надвисокої частоти, ультразвук. За допомогою ультразвукових хвиль досягається явище кавітації. На мікроорганізми діють гідравлічні удари, іонізується вода, змінюються хімічні властивості деяких речовин і вони згубно діють на клітину.
Радіохвилі – це електромагнітні хвилі, які характерізуються відносно великою довжиною – від міліметрів до кілометрів. Проходження короткочасних і ультракоротких радіохвиль через середовище індикує в ньому виникнення перемінних струмів високої та надвисокої частот. Поглинута розташованим в електромагнітному полі об’єктом електрична енергія перетворюється на теплову. Об’єкт нагрівається швидко і рівномірно по всій масі до високих температур. Загибель мікроорганізмів в електромагнітному полі високої інтенсивності настає внаслідок теплового ефекту.
Ультразвук
Ультразвук – це високочастотні коливання звукових хвиль. Ультразвук чинить могутню бактерицидну дію на прокаріоти. Сила цієї дії залежить від частоти коливань, тривалості впливу, а також від фізіологічного стану й індивідуальних особливостей мікроорганізму. При тривалому озвучуванні мікробної культури спостерігається 100% летальний ефект.
Механізм бактерицидної дії ультразвуку полягає в незворотних фізико-хімічних змінах життєво важливих компонентів мікробної клітини і механічних ушкодженнях усіх клітинних структур, що їх призводить електрогідравлічний ефект ультразвуку.
Хімічні фактори
Серед хімічних факторів розрізняють три группи:
реакція середовища – рН;
окиснювально-відновні умови;
антисептики.
Реакція середовища рН
Реакція середовища рН визначається концентрацією іонів Н і іонів ОН у водному розчині. Для кількісної характеристики реакції середовища вводять величину рН – водневий показник, який характеризує ступінь кислотності ( 7:1) або лужності (7:14). рН впливає на транспорт поживних речовин, на їх концентрацію всередині, на активність ферментів, денатурацію білків. В залежності від відношення до діапазону рН мікроорганізми класифікують на:
нейтрофіли – для них оптимальне значення рН 6,5...7,5;
ацидофіли і кислотно толерантні - оптимальне рН нижче 5;
алкалофіли – рН вище 7.
Життєдіяльність кожного виду мікроорганізмів можлива при інших сприятливих умовах лише в більш-менш визначених границях рН середовища, вище і нижче яких вона пригнічується.
Але виявляється загальна тенденція: для бактерій кисле середовище згубніше лужного, особливо для гнильних і збудників харчових отруєнь. Вегетативні клітини звичайно менш стійкі, ніж спори.
Реакція середовища впливає на розчинність речовин поживного субстрату і надходження їх у клітину. Зміна реакції середовища нерідко супроводжується підвищенням концентрації токсичних для мікробних клітин сполук. При зсуві рН у кислу сторону і підвищенні температури спостерігається різке збільшення швидкості денатурації білків у клітинах мікроорганізмів.
Стійкість прокаріотів до кислого і лужного середовища пояснюється специфікою хімізму функціонування їх клітинних бар’єрів. Крім цього, у процесі метаболізму вони здатні видаляти з клітини іони Н чи ОН, підтримуючи внутрішньоклітинне значення рН у нейтральному діапазоні.
Окислювально-відновні умови
Мікроорганізми розрізняють по потребах в умовах аерації. Тому їх класифікують за типами дихання: на аеробів та анаеробів. Аеробні клітини окислюють субстрати з виділенням енергії за реакцією:
С6 Н12 О6 + 6О2 = 6СО2 + 6Н2 О +Q
Енергія, яка вивільнюється застосовується бактеріями на біосинтез, механічну роботу і виділення енергії у вигляді світла і тепла. Теплова енергія, яка вивільнюється при диханні викликає явище термогінез, він має місце при самозігріванні або самозапалюванні зерна. Мікроби, які перетворюють хімічну енергію в світло називають фотогенними, а процес фотогенез. Анаероби не можуть застосовувати атмосферний кисень для отримання енергії вони використовують анаеробні дегідрогенази. Цей принцип відкрив Пастер і дав назву бродіння: спиртове, маслянокисле. Розрізняють підтипи облігатні та факультативні анаероби. Облігатні анаероби в лабораторіях визначаються за допомогою середовища Кітт-Тароцці. Факультативних анаеробів не виявлено.
Окисно-відновний потенціал середовища впливає не тільки на ріст і розмноження аеробних і анаеробних мікроорганізмів, але і на їх обмін речовин. Регулюючи окисно-відновні умови середовища, можна загальмувати або викликати активний розвиток тієї чи іншої групи мікроорганізмів. Можливо, наприклад, викликати ріст анаеробів у присутності повітря, додаванням речовин, які редукують та знижують окисно-відновний потенціал середовища. І навпаки, можна культивувати аероби в анаеробних умовах, підвищити рН середовища за рахунок введення речовин, які мають окисні властивості.
Антисептики
Антисептики – це хімічні сполуки, які згубно діють на мікроорганізми, їх дія зветься бактеріостатичною, коли затримується ріст та розмноження і бактерицидна – тобто загибель бактерій. Ефект дії антисептиків залежить від хімічної природи, концентрації часу експозиції та ін. факторів.
Класифікація антисептиків:
неорганічні солі – солі важких металів. Механізм дії – денатурація білка, інактивація ферментів.
окислювачі – хлор, хлорамін, хлорне вапно, спирт, розчин йоду, перманганат калію, перекис водню. Механізм дії – хлор та озон призводять до накопичення вільних активних радикалів. Формальдегід викликає денатурацію білка. Хлорамін використовують для дезінфекції в медицині, на харчових підприємствах, хлор використовують для знезараження питної води.
органічні сполуки – 70% розчин етанолу, фенол, крезол, формальдегід, поверхнево активні речовини. Механізм дії для більшості – розчинення ліпідів мембран.
Механізм дії антисептиків різний. Більшість з них порушують проникність цитоплазматичної мембрани, а після проникнення в клітину вступають у взаємодію з її компонентами, в результаті чого значно порушуються життєві процеси. Солі важких металів, формалін, феноли викликають коагуляцію білкових речовин клітини і є ферментними отрутами. Спирти, ефіри розчиняють ліпіди клітинних мембран. Хлор і озон викликають активні окисні процеси, не властиві матаболізму мікробної клітини, а також руйнують ферментні системи.
Оскільки чутливість різних мікроорганізмів до одного хімічного консерванту неоднакова, доцільне застосування їх суміші, підбираючи її склад з таким розрахунком, щоб один консервант доповнював інший у відношенні впливу на мікробі оту продукту.
Біологічні фактори
У природному середовищі існування в процесі еволюції сформувалися дуже складні і різноманітні взаємини між певними видами і групами мікроорганізмів. Серед різних ценозів значне місце в природних умовах посідають мікробіоценози – спільність мікроорганізмів. Спільне існування різних організмів одержало назву симбіозу. Симбіотичні відношення можна підрозділити на конкурентні та асоціативні (взаємно сприятливі). Асоціативні взаємовідносини мікроорганізмів у природі зустрічаються дуже часто. Серед них можна виділити декілька основних видів.
Симбіоз визначає тісні асоціативні взаємини мікроорганізмів, які стимулюють і підтримують розвиток один одного. Симбіонти спільно завжди краще розвиваються краще, ніж кожний з них окремо.
Мутуалізм – це симбіоз, при якому обидва партнери мають користь від взаємного існування. Молочнокислі бактерії, утворюючи молочну кислоту, підкисляють середовище і тим самим стимулюють розвиток дріжджів. Останні синтезують амінокислоти і вітаміни, необхідні для харчування молочнокислих бактерій.
Синергізм як різновид симбіозу характеризується тим, що при розвитку асоціації мікроорганізмів їх основні фізіологічні функції виявляються сильніше, ніж при розвитку кожного окремо.
Коменсалізм – різновид асоціативних взаємин, який передбачає стимуляцію одного мікроорганізму іншим. Так, дріжджові гриби і сарцини, що продукують різні амінокислоти і вітаміни, дуже часто сприяють росту і розмноженню інших, більш вимогливих до живильного субстрату бактерій – молочнокислих або оцтовокислих.
Метабіоз – взаємини, при яких один мікроорганізм своєю діяльністю створює умови для розвитку іншого, який як правило, продовжує процес, початий першим.
Антагонізм. Конкурентні взаємини мікроорганізмів виявляються в різній формі і виникають внаслідок різних причин. Це насамперед відносини антагонізму, при якому один мікроорганізм своєю життєдіяльністю придушує розвиток іншого. Антагоністичні відносини між мікроорганізмами виникають у боротьбі за речовини живильного середовища, за використання молекулярного кисню, за екологічну нішу і зустрічаються в природі всюди: в грунті, водоймах, в організмах тварин і людин.
Відкриття явища антагонізму належить Л.Пастеру (1877), який дослідив загибель збудника сибірської виразки в спільній культурі його з синьо гнильною паличкою. Ідею використання антагонізму мікробів в медицині вперше запропонував І.І.Мечніков. Він посилено пропагував використання молочнокислих бактерій для нормалізації мікрофлори кишечнику і придушення гнильних мікробів.
Паразитизм є крайнім проявом конкурентних взаємин мікроорганізмів. При паразитизмі один мікроорганізм використовує інший як джерело живильного субстрату, що нерідко супроводжується загибеллю жертви.
Антибіоз – різновид активних антагоністичних відносин, в основі якого лежить виділення одним мікроорганізмом речовини, токсичної для іншого. Саме на цій властивості мікроорганізмів засноване застосування антибіотиків – високоактивних речовин біологічного походження, які здатні навіть у низьких концентраціях придушувати ріст багатьох мікроорганізмів.
Антибіотики володіють виборчою здатністю пригнічувати та затримувати ріст мікроорганізмів. Кожний антибіотик характеризується специфічним спектром дії – він активний відносно певних мікроорганізмів. Деякі антибіотики впливають на гриби, інші – на бактерії. Є антибіотики, що активні по відношенню до грам негативних або, навпаки, до грам позитивних бактерій. Проте існують антибіотики з дуже широким спектром дії, яка поширюється і на гриби, і на різноманітні бактерії.
Інфекція
Інфекція – цей термін походить від латинського infectio – зараження. Інфекційний процес – це процес проникнення, розмноження патогенів в макроорганізмі. Патогени спричиняють шкоду макроорганізму і існують за його рахунок. Інфекційна хвороба – це крайній ступінь прояву інфекційного процесу. З біологічної точки зору, інфекційний процес є різновидом паразитизму. Хвороботворні мікроорганізми характеризуються наступними властивостями:
патогенність – це видова ознака характерна для певного виду, принципіальна здатність певних видів мікроорганізмів спричиняти захворювання.
специфічність – це коли окремі види патогенів можуть спричиняти лише певне захворювання.
вірулентність – це ступінь патогенності культури, окремого штаму, здатність мікроорганізмів проникати в органи і тканини, розмножуватись в них, продукувати такі речовини, які можуть послабити захисні сили макроорганізму. Вірулентність можна підвищити або послабити штучно за допомогою фізичних, хімічних і біологічних факторів. Штучне послаблення вірулентності штаму використовується при виготовленні вакцин.
За одиницю вірулентності враховують летальну дозу disis letalis minima – DLM. Летальна доза – це найменша кількість клітин патогенів, що викликає загибель сприйнятливого організму.
Одним з основних факторів патогенності є токсигеноутворення – це властивість продукувати токсигенні речовини. Мікроорганізми, які виробляють токсини називаються токсигенними. Токсини поділяються на ендо- і екзотоксини.
Екзотоксини виділяються із клітини патогенів у навколишнє середовище. До екзотоксигенів належать токсини збудників анаеробної інфекції, чуми, холери, правца. Механізм дії токсигенів полягає в тому, що вони вибірково діють на органи і тканини. За хімічною природою вони являються білками або ферментами і є термостабільними. Наприклад, токсин C.botulinum ін активується після теплової обробки протягом 12 хвилин при температурі вище 100 ºС. Людина гине від дози токсину C.botulinum 10^-8 мг.
Ендотоксини міцно зв’язані з клітинними структурами патогенів і виділяються в середовище лише після загибелі патогенна. Їх токсична активність менша ніж токсичність екзотоксинів, тому вони уражають у великих дозах. Вони також термостійкі і за хімічною природою вони полісахариди. Ендотоксини утворює кишкова паличка, сальмонелла, збудник черевного тифу.
Патогенність і вірулентність обумовлюють три фактори:
адгезивність
інвазивність
капсулоутворення.
Адгезивність обумовлена здатністю патогенів прикріплюватись до клітини епітелію органів, тканин. Реалізується ця здатність за допомогою рецепторів хемотаксису і білковими антигенами.
Інвазивність – це здатність патогенів до проникнення і розповсюдження у макроорганізмах. Для подолання бар’єрів хазяїна Патогени продукують специальні речовини, їх загальна назва – агресини. Вони з хімічною природою являють собою комплекси полі пептидів і ферментів. Ці речовини руйнують тканини макроорганізму. Наприклад, гіалуроназа, ліцитіназа, колагеназа.
Капсулоутворення. Ця ознака не є видовою, тому що у одного вида бактерій бувають капсульні і безкапсульні штами. Встановлено, що вірулентність вища у капсульних штамів.
Періоди розвитку інфекційного процесу
Розрізняють чотири періоди розвитку інфекційного процесу:
інкубаційний період. Він триває з моменту проникнення патогенна в макроорганізм до появи перших ознак захворювання. Під час першого періоду відбувається розмноження, накопичення клітин патогенів та їх токсинів. Термів цього періоду вимірюється годинами, а іноді – роками.
продромальний період. На цьому етапі відсутні характерні симптоми, розвиваються неспецифічні спільні для багатьох хвороб ознаки. Наприклад, слабкість, нездужання, втрата апетиту. Цей період іноді не буває, а може тривати до трьох днів.
в медицині цей період звуть клініка або період основних проявив захворювання. Це гарячка, запалення, ураження центральної нервової системи, порушення функції систем і органів.
видужання – реконвалесценція. Для цього періода характерно згасання клінічних проявів, зниження інтенсивності патологічних процесів. Він відбувається протягом кількох діб. Наприклад, віспа або тиждень – гепатит, бруцельоз. Коли клінічне видужання не збігається за часом зі звільненням від патогенів, вони можуть співіснувати з макроорганізмом, виділятися довгий час. Людина є носієм патогенів без проявів ознак хвороби, такий тип співіснування називається бацилоносіство.
Джерела та механізми передачі інфекції, вхідні ворота
Між трьома факторами спостерігається взаємозв’язок. Термін вхідні ворота інфекції позначає місце проникнення патогенів у макроорганізм.
| Джерела і фактори | Механізми | Вхідні ворота | Хвороба |
| Хвора людина,тварина, Речі хворого | контактний | Покривні тканини, шкіра | Сибірка, віспа, бруцельоз |
| Хвора людина, кровососні комахи | трансмісія | Покривні тканини, шкіра | Малярія, менінгіт, тиф |
| Хвора людина, повітря, грунт | аспіраційний | Дихальні шляхи | Грип, скарлатина, туберкульоз |
| Хвора юдина, питна вода, харчові продукти | аліментарний | Харчова система | Тиф, дизентерія |
Основним джерелом є хвора людина, тварина, у всіх інших випадках інфекційні хвороби звуться зооантропонози.
Класифікація інфекційних захворювань за характеристикою розповсюдження
спорадичні – окремі випадки протягом короткого часу;
епідемія – інфекція розповсюджується на значній території;
пандемія – інфекція охоплює цілі країни і континенти.
Форми інфекції
В залежності від терміну інфекційного процесу розрізняють: молнієносні – за кілька годин; гострі – за кілька діб; підгострі – за кілька тижнів; хронічні – на протязі місяців і років.
Септицемія – інфекційний процес при якому мікроорганізми розмножуються і нагромаджються в крові, проникають в усі органи і тканини організму.
Бактеріємія – інфекція, за якої кров є переносником збудника захворювання, а його розмноження відбувається в інших тканинах.
Токсенія – інфекційний процес, що викликаний збудником, який утворює токсини, які попадають у кровоток.
Імунітет
Imunitas – вільний від чого-небудь.
Імунітет – це несприятливість до всіх генетично чужорідних агентів – антигенів. Це реакції боротьби за збереження специфічності та сталості внутрішнього середовища організма. Імунітет – захист від зовнішньої чи внутрішньої біологічної агресії, захист від антигенів.
Термін „антиген” походить від грецького anti – проти та genos – рід.
Для організму роль антигенів можуть виконувати різноманітні агенти з чужорідною генетичною інформацією. Але вони завжди викликають в макроорганізмі реакції імунітету.
До антигенів відносяться:
патогенні хвороботворні бактерії;
токсини патогенних хвороботворних бактерій;
специфічні високомолекулярні речовини – білки, ферменти, полісахариди, ліпопротеїди;
власні перероджені мутантні і пухлинні клітини;
отрути рослинного і тваринного походження: змій, павуків, скорпіонів, бджіл;
гриби, віруси, рикетсії;
анатомічні структури бактерії – капсули, спори, джгутики, клітинна стінка.
Імунітет може бути таких видів: тканинний, гуморальний, функціональний.
Функціональний імунітет – це неспецифічні реакції несприятливості. Наприклад, підвищення температури, порушення діяльності центральної нервової системи.
Тканинний імунітет – це форма несприятливості, яка включає захисні властивості і бар’єрні форми тканин, клітин, організмів. Серед них можна назвати тімус – загрудинна залоза внутрішньої секреції; кістковий мозок, селезінка, лімфатичні вузли, слизові оболонки, епітеліальні шари шкіри, форменні елементи крові (лейкоцити, лімфоцити) та лімфоїдні фолікули кішок. Всі вони здатні до фагоцитозу. Фагоцитарна теорія імунітету була розроблена І.І.Мечниковим. Фагоцитоз – це внутрішньоклітинне травлення. Процес фагоцитозу йде по фазам:
наближення фагоцита до патогенна;
адсорбція і поглинання мікробу у фагосому;
утворення фаголізосоми;
ферментативне перетравлення.
Фагоцити – це клітини, які поглинають і перетравлюють живих і мертив мікробів, а також інших генетично чужорідних агентів.
Гуморальний імунітет – це несприятливість, яка зв’язана з бактерицидними властивостями крові і лімфи. Розрізняють неспецифічні фактори гуморального імунітету (комплімент, лейкіни) і специфічні – антитіла.
Антитіла – це високо специфічні білки, які утворюються і накопичуються в крові при введенні антигенів і вступають з ними у взаємодію. Розрізняють кілька класів імуноглобулінів за критеріями – молекулярна маса, первинна структура білка, фізико-хімічна структура, функції, концентрація в корі, антигенність. За механізмом дії їх класифікують:
аглютеніти – антитіла, які спричиняють склеювання антигена з утворенням конгломератів. Ця реакція зветься реакцією аглютинації і її умовно позначають:
RA = An + Ag
В 1895р. Вчений Відаль застосовує сироватку крові з антитілами для діагностики черевного тифу. І зараз цю реакцію використовують для визначення виду бактерій. Наприклад, для ідентифікації кишкової палички, сальмонели, для діагностики різних захворювань.
преципітини – антитіла, які утворюють з антигеном дрібний осад – преципітат, а реакція називається реакцією преципітації, її застосовують для діагностики сибірки.
антитоксини – антитіла, які нейтралізують токсини.
лізини – антитіла, які утворюються в сироваткі крові і проводять лізис антигенів.
Імунітет розрізняють слідуючи видів: імунітет природжений або спадковий. Це несприятливість до інфекції, яка передається від матері до дитини. Якщо несприятливість викликає в процесі життєдіяльності, то імунітет зветься набутий. Він поділяється на активний і пасивний.
Пасивний набутий імунітет створюється введенням в організм сироватки з готовими антитілами проти збудника хвороби, тобто проти антигена. Сироватку добувають у донора, який переніс відому хворобу.
Активний набутий імунітет – це результат перенесення хвороби, або вакцинації. Вакцина – це препарат із живих збудників захворювання із послабленою вірулентністю або з убитих патогенних. Вакцини поділяються на чотири групи за складом:
вакцини з живих патогенів з послабленою вірулентністю – це сибірка, сказ, грип, чума.
убиті збудники хвороби – від холери, тифа.
вакцини, які складаються з антитоксинів. Антитоксини виробляють шляхом обробки екзотоксинів формаліном або іншими шляхами – дифтерія холера, ботулізм.
вакцини, які являють собою комплексні препарати, хімічні речовини з авірулентних або послаблених вірулентних токсинів.
У формуванні природного імунітету важливу роль відіграє нормальна мікрофлора в організмі людини і тварин. Облігатні мікроорганізми, які продукують речовини типу антибіотиків, молочну кислоту, спирти, пероксид водню та інші сполуки, проявляють антагоністичні властивості відносно багатьох патогенних бактерій. Якісні і кількісні порушення в складі мікро біоценозів у організмі людини одержали назву дисбактеріозів. Дисбактеріози виникають найчастіше в результаті тривалого прийому антибіотиків, а також при хронічних інфекціях, внаслідок дії радіації і дії екстремальних факторів. При дисбактеріозі відбувається придушення облігатної мікрофлори мікроорганізму і активне розмноження умовно-патогенних бактерій (Proteus, Pseudomonas) і дріжджових грибів Candida albicans.
Аліментарні (харчові) захворювання
Захворювання, які викликають в результаті вживання харчових продуктів називають аліментарними, харчовими або кишковими захворюваннями. Найчастіше їх спричиняють представники родини ентеробактерій. До цієї родини входять патогенні та умовно патогенні мікроорганізми.
Харчові захворювання поділяються на харчові інфекції та харчові отруєння. Харчові отруєння поділяються на дві підгрупи: інтоксикації (токсикози) та токсико-інфекції.
Харчові інфекції
Харчові інфекції – заразні захворювання. Вони розповсюджуються не тільки через харчові продукти, але і через воду, повітря, контактно-побутовим та іншими шляхами. Більшість збудників цих захворювань у харчових продуктах, вони здатні довгий час зберігати життєздатність та вірулентність. Для виникнення захворювання достатньо проникнення до організму невеликої кількості збудників. Інкубаційний період таких захворювань може тривати від кількох днів до декількох тижнів.
Потрапивши до організму людини більшість збудників харчових інфекцій локалізується головним чином у кишечнику, тому їх називають також кишковими інфекціями. Найбільшу небезпеку представляють такі кишкові інфекції, як черевний тиф, паратифи, дизентерія, холера та вірусний гепатит. Такі харчові інфекції як бруцельоз, туберкульоз, сибірська виразка, ящур передаються людині від тварини через травний шлях, але крім кишково-шлункового тракту вражають й інші системи організму.
Черевний тиф та паратифи А і В викликаються бактеріями роду Salmonella, які мають багато загальних властивостей. Сальмонели – це рухомі короткі грам негативні палички, факультативні анаероби, які не утворюють спор та капсул. Вони не синтезують екзотоксин, але при руйнуванні бактеріальних клітин в організмі виділяється високотоксичний термостабільний ендотоксин.
Оптимальна температура їх розвитку 37 ºС, але вони ростуть і при 15-40 ºС. При нагріванні до 60 ºС сальмонели гинуть через 30-45 хвилин. При кип’ятінні та при обробці дезінфікуючими розчинами гинуть через кілька секунд. Добре витримують низькі температури, високі концентрації солі та висушування. Знаходячись у харчових продуктах, особливо м’ясних, проявляють значну термостійкість.
При профілактиці сальмонельозів важливими є виявлення бактеріоносіїв та їх ізоляція, боротьба з мухами, які є переносниками сальмонел, додержання правил приготування та зберігання харчових продуктів.
Дизентерія (або, на відміну від дизентерії, що викликається амебами, бактеріальна дизентерія) викликається низкою близьких між собою бактерій, об’єднаних в рід Shigella. Найбільш розповсюдженими є паличка Зонне, паличка Флекснера, паличка Григоьєва-Шига. Шигели – це короткі грам негативні нерухливі палички, що не утворюють спор та капсул, факультативні анаероби, утворюють ендотоксин. Збудники дизентерії не розщеплюють сечовину, не володіють протеолітичними властивостями, не утворюють сірководню та індолу, тому при розвитку в харчових продуктах вони не змінюють їхні органолептичні властивості.
Шигели розмножуються в інтервалі температур 10-45 ºС, оптимальна температура – 37 ºС. Вони гинуть при нагріванні до 60 ºС через 10-15 хвилин, у розчинах хлораміну та хлорного вапна через 20-30 хвилин. У харчових продуктах, на посуді зберігаються в середньому до 10-20 діб. В залежності від дози інфекції, рН середовища та інших умов можуть зберігатися і довше. При споживанні продуктів, що містять велику кількість збудників дизентерії, захворюванні проходять як харчове отруєння.
Профілактика захворювання передбачає своєчасне виявлення та ізоляцію хворих та бактеріоносіїв, додержання санітарно-гігієнічних правил у побуті й на підприємствах харчової промисловості та громадського харчування.
Холера відноситься до так званих особливо небезпечних інфекцій, вона викликає високу смертність серед хворих, особливо серед дітей та людей похилого віку. Збудник цього захворювання – холерний вібріон має форму зап’ятої та характеризується поліморфізмом, може утворювати пари чи короткі ланцюжки. Він рухливий, не утворює спор та капсул, грам негативний аероб. Продукує екзотоксин, ендотоксин та багато ферментів патогенності. Ферментує білки з виділенням сірководню та індолу, та вуглеводи – без утворення газоподібних продуктів.
Холерний вібріон розмножується тільки в лужному або нейтральному середовищі (рН 7,6 – 8,6) при 14-42 ºС. Він гине при нагріванні до 80ºС через 5 хвилин, при 100 ºС – миттєво. Чутливий до дії ультрафіолету, кислот, висушування.
Єдиним джерелом холерних вібріонів є людина. Захворювання може швидко розповсюджуватись через джерела водопостачання та харчові продукти і викликати епідемії. Профілактика холери передбачає дотримання звичайних санітарно-гігієнічних правил, але при загрозі виникнення епідемії використовують специфічні методи профілактики – вакцинацію протихолерною вакциною та застосування холерного бактеріофагу, який приймають через рот та додають до джерел водопостачання.
Харчові отруєння
Харчові отруєння можуть бути немікробного та мікробного походження. Причиною немікробного харчового отруєння може бути вживання отрутних грибів чи рослин, а також продуктів, що забруднені токсичноми речовинами антропогенного походження – токсичних органічних сполук або токсичних хімічних елементів і таке інще. Харчові отруєння мікробної етіології умовно поділяють на токсикоінфекції та харчові інтоксикації.
Харчові токсикоінфекції
Харчові токсикоінфекції – гострі кишкові захворювання, яки виникають в результаті вживання харчових продуктів, котрі містять велику кількість живих мікробів та їх токсинів. Інкубаційний період продовжується кілька годин, рідше більше доби. З шлунково-кишкового тракту мікроорганізми через лімфу та кров потрапляють до інших органів. Внаслідок їхньої загибелі вивільнюється ендотоксин, який викликає патологічні зміни в уражених органах та здійснює токсичний вплив на центральну нервову систему. Збудники харчових токсикоінфекцій від хворих людей до здорових не передаються.
Сальмонельозні токсикоінфекції виникають при застосуванні в їжу продуктів, які містять значну кількість бактерій роду Salmonella. Інкубаційний період захворювання короткий (декілька годин), воно супроводжується підвищенням температури, загальної слабкістю, білем у кишечнику та іншими явищами гастроентериту.
При розмноженні сальмонел у харчових продуктах їхній зовнішній вигляд та смак не змінюється. У готових виробах ці бактерії при кімнатній температурі розмножуються значно швидше, ніж у сирих. Особливої уваги потребує м’ясний фарш, оскільки подрібнення та перемішування м’яса створює сприятливі умови для розмноження мікробів. Треба також приймати до уваги, що час, який використовується для приготування страв в мікрохвильових печах, є недостатнім для знищення бактерій роду Salmonella.
Токсикоінфекції, що викликаються умовно-патогенними бактеріями. Умовно-патогенні бактерії є постійними мешканцями організму людини (шкіри, кишечнику, дихальних шляхів). За нормальних умов вони не викликають захворювань, але при ослабленні макроорганізму можуть стати причиною хвороби.
Кишкова паличка (Escherichia coli) є постійним мешканцем нижнього відділу кишечнику людей та тварин, тому виявлення типових кишкових паличок у воді, ґрунті, харчових продуктах свідчить про свіже фекальне забруднення цих об’єктів. Зараження продуктів бактеріями групи кишкової палички відбувається при порушенні санітарного режиму на виробництві, при виготовленні, зберіганні та реалізації продукції.
За морфологічними ознаками це маленькі, короткі, грам негативні, рухомі палички, що не утворюють спор та капсул. Вони є факультативними анаеробами, культиву.ться на простих середовищах при рН 7,2-7,4, та оптимальній температурі 37 ºС. Кишкові палички гинуть при звичайних методах стерилізації при 67-75ºС.
На МПА кишкова паличка утворює напівпрозорі з сірувато-блакитним відтінком круглі колонії з рівними краями. На середовищі Ендо вона утворює червоні колонії середнього розміру з металевим блиском. Кишкові палички виробляють термостабільний ендотоксин – ентеротропну отруту та термолабільний екзотоксин, що є нейротропною отрутою. У маленьких дітей та ослаблених дорослих кишечна паличка може потрапляти з нижніх відділів та інших органів і викликати тяжкі кишкові захворювання та важкі запальні процеси. Окрім умовно-патогенних типів зустрічаються такі серологічні типи кишкової палички,0 які мають властивості патогенних мікроорганізмів. Вони викликають колі ентерити у маленьких дітей.
Бактерії роду протесу (Proteus) зустрічаються у кишечнику теплокровних тварин та людей, в ґрунті, стічних водах, у білкових продуктах, що гниють. Це прямі поліморфні грам негативні палички, рухомі, з перитрихіальним розташуванням джгутиків, спор та капсул не утворюють, факультативні анаероби, за типом обміну речовин – хемоорганогетеротрофи, які мають і дихальний, і бродильний типи метаболізму. Ростуть при температурах від 5 ºС до 43 ºС, стійкі до низьких температур, а при 80 ºС гинуть за 5 хвилин. У посівах в конденсаційну воду скошеного МПА через 12-18 годин спостерігається повзучий ріст протея у вигляді вуглеподібної плівки.
Стрептококи, які відносяться до роду Streptococcus, є збудниками гнійно-запальних процесів, сепсису (зараження крові), окремі з них ентеротоксичні – можуть викликати харчові отруєння. Харчові отруєння може викликати також фекальний стафілокок, який входить до складу нормальної мікрофлори кишечника людини та тварин.
За морфологічними ознаками стрептококи – грампозитивні короткі ланцюжки коків, які не утворюють спор та капсул. Вони поліморфні: клітини відрізняються за розмірами та формою. Стрептококи погано ростуть на звичайних поживних середовищах. На МПА з глюкозою (0,5-1,0%) стрептококи утворюють сірувато-білі колонії діаметром до 1 мм; на МПБ ростуть у вигляді пластівців та осаду.
Стрептококи витримують температуру 60-65 ºС протягом 30 хвилин, здатні рости в присутності 6,5% хлориду натрію, в середовищах з рН 9,6-10. У висушеному стані можуть зберігатися протягом 4-6 місяців. Під дією прямих сонячних променів гинуть через 2-3 години, при кип’ятінні гинуть миттєво.
Харчові інтоксикації
Харчові інтоксикації пов’язані із вживанням в їжу продуктів, в яких в результаті життєдіяльності токсиноутворюючих мікроорганізмів накопичився екзотоксин. При цьому живих токсиноутворюючих мікроорганізмів може в продукті уже й не бути. Після вживання таких продуктів токсин всмоктується через шлунково-кишковий тракт у кров і розноситься по всьому організму. Токсин, у першу чергу, вражає серцево-судинну та центральну нервову системи.
Інкубаційний період при токсикозах коротший, ніж при харчових токсикоінфекціях і складає кілька годин. Збудниками харчових токсикозів є патогенні стафілококи, стрептококи, збудники ботулізму та токсигенні гриби.
Ботулізм. Термін ”ботулізм” походить від латинського слова botulus – ковбаса. Таку назву захворювання отримало, оскільки збудник ботулізму Clostridium botulinum вперше був виділений із ковбаси.
Clostridium botulinum – велика поліморфна паличка із заокругленими кінцями довжиною 4-8мкм, шириною до 13 мкм, розташовується хаотично, іноді парами або у вигляді коротких ланцюжків. Паличка рухома, перетри. Грампозитивна, капсул не утворює. Спори розташовуються в клітині субтермінально, тому клітина зі спорою нагадую ракетку або ложку. Паличка ботулізму є суворим анаеробом.
Оптимальною температурою росту та токсиноутворення є 30-40 ºС, оптимальне рН 7,2-7,4 (при рН 3,0-4,0 не розмножується). На середовищі Кітт-Тароцці бактерії створюють помутніння та осад. Бульйон набуває запаху згірклого масла.
Збудник ботулізму синтезує два головних типи екзотоксинів: нейротоксин та гемолізи. Нейротоксин є простим білком. Це найсильніша отрута з усіх відомих у світі отрут. Саме він створює клінічну картину інтоксикації при ботулізмі. Цей токсин не руйнується протеолітичними ферментами шлунково-кишкового тракту.
Ботулінічний токсин повністю ін активується в харчових продуктах при нагріванні до 80 ºС протягом 30 хвилин, а при 100 ºС – за декілько хвилин. Тому звичайна кулінарна обробка продукту повністю цого ін активує.
Спори зберігаються довгий час, а в сприятливих умовах у літній період можуть проростати та розмножуватися. Спори збудника ботулізму термостабільні – стійкі до нагрівання. При кип’ятінні зберігаються протягом 5-6 годин, при автоклаву ванні гинуть через 20-30 хвилин. Спори стійкі до багатьох бактерицидних речовин: 20% розчин формаліну вбиває їх через 24 години; етиловий спирт – через 2 місяці; 10% соляна кислота – лише через одну годину.
Зовнішні ознаки зараження консервів збудником ботулізму – бомбаж, зміна структури продукту, неприємний запах – спостерігається не завжди.
Інтоксикації грибкової природи називають мікотоксикозами, а метаболіти грибів, які мають токсичні властивості, називають мікотоксинами. Відомо більш двохсот індивідуальних мікотоксинів. Найбільш повно вивчена хімічна природа афлатоксинів, зеараленону, трихотеценів, охратоксинів, стеригматоцистину.
Більшість мікотоксинів термостійкі, канцерогенні, мають кумулятивні властивості. Мікотоксини можуть зберігатися в продуктах навіть при стерилізації.
Для запобігання ураження плісеневими грибами загальноприйняті технології зберігання сировини доповняють обробкою спеціальними хімічними препаратами – плісеневими інгібіторами, які містять органічні кислоти та їх солі.
Характеристика санітарно-показових мікроорганізмів
Санітарно-показові мікроорганізми є індикаторами при альтернативному методу визначення якості продукта і дозволяють визначити небезпечність продукта з точки зору епідемії, виникнення отруєння, а також оцінити ефективність техніки термічних обробок продукта і дати оцінку санітарно-гігієнічного стану.
Такі санітарно-показові мікроорганізми, як кишкова паличка, фекальні ентерококи, сульфітредукуючі клостридії та протесу є показниками можливої присутності в об’єктах зовнішнього середовища збудників кишкових захворювань. Наявність в об’єктах зовнішнього середовища стрептококів та стафілококів є свідченням можливості присутності збудників інфекцій дихальних шляхів.
Кишкова паличка (Escherichia coli) має найбільше значення як санітарно-показовий мікроорганізм. Бактерії роду Escherichia – це дрібні рухливі грам негативні палички, що не утворюють спор, не володіють оксидазною активністю, ферментують лактозу і глюкозу з утворенням кислоти та газу. Вони є факультативними анаеробами, добре ростуть в універсальних живильних середовищах, стійкі до дії багатьох анілінових барвників, не розріджують желатин, здатні ферментувати ряд вуглеводів – лактозу, глюкозу, мальтозу, сахарозу з утворенням кислоти і газу.
Для диференціації використовують середовище Ендо, на якому Escherichia coli дає характерний ріст у вигляді колоній червоного кольору з металевим блиском. Середовище Ендо є селективним середовищем для ентеробактерій, у його склад входять МПА, лактоза, фуксин основний, сульфат і фосфат натрію.
Виявлення бактерій роду Escherichia в харчових продуктах, у воді, ґрунті, на устаткуванні свідчить про свіже фекальне забруднення цих об’єктів. Наявність кишкової палички свідчить про ризик харчового отруєння. Виявлення кишкової палички проводять для визначення правильності проведення термічної обробки при виготовленні ковбасних виробів, пастеризації молока тощо, а також при дослідженні санітарно-гігієнічного стану виробництва, оцінці ефективності застосування дезінфектантів, миття обладнання, апаратури та рук працівників харчового виробництва. Державним стандартом і Санітарними правилами і нормами обмежено кількість кишкової палички в одиниці продукту.
Кількість кишкової палички в об’єктах зовнішнього середовища характеризують двомав показниками: колі-титром та колі-індексом. Колі-титр – це найменша кількість досліджуваного матеріалу, в якій виявлено одну клітину кишкової палички. Чим менша величина колі-титру, тим більш небезпечний даний об’єкт у епідеміологічному відношенні. Колі-індекс – це кількість клітин кишкової палички в одиниці об’єму або маси досліджуваного матеріалу.
Ентерококи поряд з бактеріями групи кишкових паличок є постійними мешканцями кишечника людини і теплокровних тварин, які у великій кількості виділяються в зовнішнє середовище. Їх виявлення у харчових продуктах, воді, ґрунті свідчить про свіже фекальне забрудненя цих об’єктів.
Ентерококи або фекальні стрептококи відносяться до роду Streptococcus. Ентерококи це грампозтивні коки, що розміщуються попарно або утворюють короткі ланцюжки. Ростуть вони при температурі від 10 до 45 ºС, оптимальна температура розвитку – 37 ºС. Для визначення ентерококів в чистій культурі з об’єктів, густо засіяних сторонньою мікрофлорою, частіше використовують селективне молочне середовище Каліни з поліміксином.
Ентерококи надзвичайно стійкі до низьких температур, нагрівання, хлорування, до підвищених концентрацій цукру і солі, до високої кислотності. В зовнішньому середовищі вони відмирають швидше, ніж кишкова паличка. У воді і ґрунті не розмножується. Тому ентерококи є показником свіжого фекального забруднення.
В Україні ентерококи поряд з бактеріями грипи кишкової палички використовують як санітарно показові мікроорганізми при санітарній оцінці води відкритих водоймищ, особливо колодязів, вода яких використовується в технологічному процесі.
Сульфітредукуючі клостридії. Серед численних патогенних і сапрофітних видів роду Clostridium як санітарно-показові використовують ті, постійним місцем перебування яких є кишечник людини і теплокровних тварин. Найбільш частим мешканцем кишечнику людини є C. perfringens. Використання C. perfringens у якості санітарно-покзового мікроорганізму ґрунтується на тому, що його спори в зовнішньому середовищі не моють високої стійкості, у харчових продуктах він розмножується тільки при температурі 18-20 ºС і вище, через 6-8 годин зберігання продукту його розмноження сповільнюється, а згодом зовсім припиняється. Особливо чутливий C. perfringens до кислої реакції середовища.
Оскільки тільки клостридії кишкового надходження мають редукуючи (відновлюючі) властивості при рості на залізосульфітних середовищах і утворюють на них колонії чорного кольору, ця ознака є основною для судження про санітарну показовість таких мікроорганізмів.
Визначення сульфітредукуючихк клостридій здійснюється шляхом висіву визначеної кількості продукту і його розведень у пробірки з напіврідким селективно-діагностичним сульфіт-залізним середовищем та культивуванні посіві при 37 ºС протяго 24-72 годин. При обліку результатів визначають мінімальну кількість продукту, у якому виявлені грампозитивні нерухомі палички, що утворюють яйцеподібні або кулясті спори та редукують сульфіт.
Бактерії роду Proteus широко розповсюджені в природі, в основному в місцях, де відбувається аеробні процеси гнильного розпаду. З кишечника ці мікроорганізми виділяються у 5-10% здорових людей. У ви породженнях коней, рогатої худоби, гризунів і інших тварин присутність паличок протея більш часта.
Температурний діапазон росту бактерій грипи протея лежить у межах 10-40 ºС. Вони нетермостійки – гинуть при прийнятих у промисловості режимах пастеризації, але стійкі до заморожування, дії антибіотиків і хімічних препаратів.
Бактерії групи протея можуть розвиватися в харчових продуктах, що містять білки. Чисті культури протея при рясному розмноження протягом 2-3 діб можуть не викликати змін органолептичних властивостей, зовнішні ознаки гниття з’являться лише в результаті спільної дії протея і спороутворюючих гнильних аеробів.
Група протея не має самостійного значення як показник фекального забруднення. Однак її присутність у великих кількостях в харчових продуктах свідчить про наявність розкладу білка. При вмісті 1-2 млрд. клітин у 1 см³ харчові продукти небезпечні для вживання і можуть служити причиною харчового отруєння внаслідок виділення з загиблих клітин протея ендотоксину з ентеротропною дією і гемолізи ну.
Стафілококи як санітарно-показові мікроорганізми часто використовуют при оцінці стану повітря, гігієнічного стану лікувальних і дитячих установ, де є небезпечним розвиток патогенних стафілококів. Для харчових продуктів цей показник має інше значення, тому що занепокоєння викликає не патогенність стафілококів, а їхня здатність виробляти ентеротоксин, що обумовлює харчові інтоксикації.
З метою санітарно-гігієнічної оцінки продукти досліджують на наявність Staphylococcus aureus. Методи виявлення стафілококів засновані на їх здатності рости на середовищах з підвищеним вмістом кухарської солі. Для виділення культур токсигенних стафілококів і з метою диференціації їх від сапрофітів частіше використовують елективно-диференціальні живільні середовища – жовточно-сольовий агар (ЖСА) і молочно-сольовий агар (МСА). Поряд з цим перевіряють наявність гемотоксину на середовищі Цейслера, наявність плазмокоагулази, дезоксирибонуклеази, здійснюють фагодіагностику, імунодіагностику (використовують реакцію преципітації), проводять біопробу.
Дріжджі та плісені часто є збудниками псування молочних продуктів, у зв’язку з цим їх називають ”технічно шкідливими” мікроорганізмами. Серед них присутні і патогенні представники, що можуть викликити мікози і мікотоксикози. Основний вид псування продуктів, що вони викликають – ”пліснявіння”.
Плісняві гриби, потрапляючи в продукт і розвиваються гнильні бактерії. Дріжджі і плісені псують товарний вид продуктів, викликають гідроліз жиру з утворенням жирних кислот, що веде до згіркнення продуктів.
Плісені викликають гнильне псування продуктів і обумовлюють їх гіркий смак. Дріжджі обумовлюють ослизнення, що скорочує термін зберігання продуктів в охолодженому стані. Характерною особливістю дріжджів є їх здатність розвиватися в середовищах, що містять до 24% NaCl і до 60% сахарози.
Спори плісеневих грибів постійно живуть у повітрі, ґрунті, гної, у продуктах, на поверхні різних предметів, стінах сирих приміщень та ін. Молочна плісень переважно живе в молочних продуктах. Плісені дуже стійкі до низьких температур. Як психрофіли, деякі плісені родів Thamnidium, Rhizopus і Cladosporium можуть розвиватися в холодильниках при 9-11 ºС.
При посіві повітря цехових приміщень седиментаційним методом протягом 5 хвилин на щільному живильному середовищі повинно виявлятися не більше 15 колоній плісеней і 19 колоній дріжджів.
Похожие работы

... - президент Української Академії Наук, а з 1929 року - дійсний член Академії Наук СРСР. В Києві заснував Інститут мікробіології та епідеміології і був його директором. Д.К. Заболотний помер від важкої хвороби 15 грудня 1929 року в м. Києві. Поховали Данила Кириловича Заболотного в його рідному селі Чоботарка. Будинок де жив учений перетворений в Музей-садибу Д.К. Заболотного, а село Чоботарка ...
... ігання маринованої риби в герметично закритій тарі і на холоді запобігає її пліснявінню. Висушування риби і в'ялення - давні способи її збереження як харчового продукту. При видаленні з риби води до певної межі створюються несприятливі умови для розвитку мікробів. Консервуючу дію у в'яленій і солено-сушеній рибі надає також сіль. Деякі мікроорганізми тривало зберігаються на цій рибній продукції ...


... результатів. Анотація Куцик Р.В. Мікробіологічне обгрунтування нових підходів до лікування та профілактики стафілококових інфекцій на основі дослідження протимікробних властивостей похідних тіазолідину, фурану, хіноліну, акридину і біологічно активних речовин природного походження. – Рукопис. Дисертацiя на здобуття наукового ступеню доктора медичних наук за спеціальністю 03.00.07 – мі ...
... і співдоповідей, розподілити завдання, повідомити питання, що будуть обговорюватися на семінарі. Крім тих методів опитування, що практикуються на основних предметах біологічного циклу, на факультативних заняттях можна застосовувати залікові заняття, що дозволяють забезпечити фронтальну перевірку всіх учнів факультативної групи. Ці заняття звичайно проводяться після вивчення чергової теми. Вони ...
0 комментариев