Навигация
Методы и условия культивирования изолированных клеток и тканей растений
39148
знаков
0
таблиц
0
изображений
Автономное образовательное учреждение
высшего профессионального образования
Ленинградский Государственный Университет
имени А.С. Пушкина
Методы и условия культивирования изолированных клеток и тканей растений
Санкт-Петербург
2009
Содержание
Введение
1.Вспомогательное использование методов in vitro в селекции растений
2. Клеточная селекция растений
3. Гибридизация соматических клеток
Литература
Введение
Одно из направлений клеточных технологий – это использование их в селекции, которое облегчает и ускоряет традиционный селекционный процесс в создании новых форм и сортов растений. Существующие методы культивирования изолированных клеток и тканей in vitro условно можно разделить на две группы.
Первая группа – это вспомогательные технологии, которые не подменяют обычную селекцию, а служат ей. К ним можно отнести: оплодотворение in vitro (преодоление программной несовместимости), культивирование семяпочек и незрелых гибридных зародышей (преодоление постгамной несовместимости), получение гаплоидов путем культивирования пыльников и микроспор, криосохранение изолированных клеток, тканей и органов, клональное микроразмножение отдаленных гибридов.
Вторая группа методов ведет к самостоятельному, независимому от традиционных методов селекции, получению новых форм и сортов растений: клеточная селекция с использованием каллусной ткани, соматическая гибридизация (слияние изолированных протопластов и получение неполовых гибридов), применение методов генной инженерии.
1. Вспомогательное использование методов in vitro в селекции растений
В отдаленной гибридизации находят применение такие методы культуры изолированных тканей, как оплодотворение in vitro, эмбриокультура (выращивание изолированных зародышей на искусственных питательных средах), клональное микроразмножение ценных гибридов, а также получение гаплоидов in vitro и криосохранение.
Оплодотворение in vitro (преодоление прогамной несовместимости) проводится в том случае, когда невозможно осуществить оплодотворение между выбранными парами в естественных условиях. Это вызвано несколькими причинами: 1) физиологические (несоответствие во времени созревания пыльцы и т.д.); 2) морфологические (короткая пыльцевая трубка или блокирование роста ее на разных этапах развития и т.д.). Оплодотворение in vitro можно осуществить двумя способами: а) культивирование на искусственной агаризованной питательной среде завязи с нанесенной на нее готовой пыльцой; б) завязь вскрывается и на питательную среду переносятся кусочки плаценты с семяпочками, вблизи которых или непосредственно на ткани плаценты культивируется готовая пыльца. Визуально определить, прошло оплодотворение in vitro или нет, можно по быстроувеличивающимся в размерах семяпочкам. Сформировавшийся зародыш, как правило, не переходит в состояние покоя, а сразу прорастает и дает начало гибридному поколению. Плацентарное оплодотворение in vitro позволило преодолеть несовместимость в скрещивании сортов культурного табака N. tabacum с дикими видами N. rosulata и N. debneyi и сделало возможным получение межвидовых гибридов табака в опытах М.Ф. Терновского и др. (1976), Шинкаревой (1986).
Преодоление постгамной несовместимости. Постгамная несовместимость при отдаленной гибридизации возникает после оплодотворения. Часто при этом образуются щуплые невсхожие семена. Причиной может быть расхождение во времени развития зародыша и эндосперма. Из-за слабого развития эндосперма зародыш бывает неспособен к нормальному прорастанию. В таких случаях из зрелой щуплой зерновки изолируют зародыш и выращивают его в питательной среде.
Выращивание зародышей в искусственной питательной среде называется эмбриокультурой. Среда для выращивания зрелого зародыша может быть простой, без добавок физиологичеки активных веществ (например, среда Уайта) или любая другая, содержащая минеральные соли и сахарозу. При более отдаленных скрещиваниях нарушения в развитии зародыша могут наблюдаться уже на ранних этапах, что выражается в отсутствии дифференцировки, замедленном росте. В этом случае культура зародыша состоит из двух этапов – эмбрионального роста зародыша, во время которого продолжается его дифференцировка, и прорастания подросшего зародыша. Для первого этапа требуется более сложная по составу среда с повышенным содержанием сахарозы, с добавками различных аминокислот, витаминов и гормонов.
Применение эмбриокультуры в селекции приобретает в последнее время большое значение для получения отдаленных гибридов зерновых, злаковых и других сельскохозяйственных культур. Показана возможность увеличения выхода пшенично-ржаных гибридов путем доращивания незрелых зародышей, а также использования эмбриокультуры для преодоления постгамной несовместимости при гибридизации пшеницы с колосняком.
Метод эмбриокультуры находит все более широкое применение в межвидовой гибридизации овощных растений. Для лука разработаны приемы выращивания in vitro абортивных зародышей от гибридных семян с разных этапов эмбриогенеза, выращивание зародышей от частично фертильных межвидовых гибридов. Культура изолированных зародышей используется в селекции томатов и других овощных растений.
Исследована гормональная регуляция роста и развития зародышей томата in vitro. Обсуждается возможность применения эмбриокультуры для получения отдаленных гибридов подсолнечника, изучаются факторы, контролирующие рост и развитие in vitro зародышей подсолнечника, выделенных в разные сроки после опыления.
Культура изолированных зародышей как вспомогательный метод при отдаленной гибридизации применяется не только для преодоления постгамной несовместимости, но также с целью микроразмножения ценных гибридов. В этом случае микроразмножение идет путем каллусогенеза, индукции морфогенеза и получения растений-регенерантов из каллусной ткани. Техника клонирования незрелых зародышей позволяет размножать ценные генотипы растений на ранних стадиях жизненного цикла. Еще одна возможность применения культуры зародышей–использование ее в клеточной селекции.
Клональное микроразмножение отдаленных гибридов. Эмбриокультура дает возможность вырастить гибридные растения из неполноценных зародышей. Однако выход гибридных растений мал, и гибриды часто бывают стерильны. Иногда, например, при селекции гречихи, трудно воспроизвести в потомстве уникальные генотипы из-за перекрестного опыления культуры. Поэтому перед исследователями часто встает задача – размножить и сохранить полученные растения. В этом помогает метод клонального микроразмножения. Размножают гибриды путем активации развития меристемы пазушных почек (черенкованием стерильных побегов), адвентивными почками или регенерацией растений из каллусной ткани, в частности полученной при культивировании зародышей.
Получение гаплоидов in vitro и использование их в селекции. Роль гаплоидных растений в селекции очень велика. Применение их позволяет быстрее найти нужную комбинацию, сокращает время для создания сорта. Гаплоиды используются для получения стабильных гомозиготных линий. Для мутагенеза также удобнее использовать гаплоиды, поскольку на гаплоидном уровне облегчается отбор рецессивных мутаций.
В диплоидных растениях мутации редко затрагивают оба аллельных гена в гомологичных хромосомах. Особь обычно гетерозиготна (два гена различаются), при этом проявляется действие только доминантного (но не рецессивного) гена. Поскольку мутации чаще рецессивны, чем доминантны, их довольно сложно выявить. В гаплоидных же растениях, которые содержат только одну из каждой пары гомологичных хромосом, мутации проявляются немедленно. Селекция на гаплоидном уровне позволяет вести прямой отбор не только доминантных, но и рецессивных признаков.
Гаплоидные особи стерильны, но можно искусственно удвоить набор их хромосом с помощью колхицина и получить диплоидные гомозиготные растения.
Гаплоиды могут возникать спонтанно, но частота их спонтанного возникновения очень мала. Искусственным путем с Использованием методов in vitro удается получить большие количества гаплоидных растений. Существует три способа получения гаплоидов с использованием метода культуры изолированных тканей:
андрогенез – получение гаплоидных растений на искусственной питательной среде из изолированных пыльников и микроспор.
гиногенез – получение гаплоидных растений на искусственной питательной среде из изолированных семяпочек;
партеногенез – получение гаплоидов из гибридного зародыша, у которого из-за несовместимости хромосом родителей потеряны отцовские хромосомы.
Образовавшиеся в результате элиминации хромосом отцовского генома гаплоидные эмбриоиды культивируют на искусственных питательных средах и получают гаплоидные растения. Сорта ячменя Исток и Одесская-15 были получены комбинацией партеногенетического метода с культурой изолированных зародышей за четыре года вместо обычных 10–12 лет. Методом культуры пыльников из сортов и гибридов мягкой и твердой пшеницы в НПО «Элита Поволжья» за четыре года получено более 2,5 тыс. дигаплоидных линий, которые характеризуются гомогенностью и стабильностью.
Продолжается разработка технологии получения гаплоидов посредством культуры пыльников пшеницы, ячменя, кукурузы, озимой ржи, картофеля. В культуре пыльников возможны два пути образования гаплоидных растений. Первый – образование растений путем эмбриогенеза в пыльцевых зернах. При этом внутри пыльников из отдельных пыльцевых зерен возникают эмбриоиды. Они прорастают и дают гаплоидные растения. Второй – образование каллуса из клеток пыльника. В дальнейшем в результате морфогенеза из каллусных клеток регенерируют растения. В этом случае образовавшиеся растения не всегда бывают гаплоидными и часто отличаются по плоидности. До конца не выяснено, образуются ли они от полиплоидизированных гаплоидных клеток или от слившихся клеток.
Гаплоиды, полученные in vitro, могут применяться не только в практической селекции, но и в работах по генетической инженерии, а также по клеточной селекции. Пыльцевые зерна являются в некоторых случаях более удобными, чем протопласты, объектами для опытов по генетической трансформации.
Криосохранение растений. Криосохранение соматических клеток растений в жидком азоте (температура – 196° С) – новое направление в биотехнологии, которое широко стало развиваться с начала 70-х годов XX столетия. Цель данной технологии заключается в сохранении в культуре in vitro генофонда, а также в обеспечении селекционеров в любое время генотипом, имеющим искомые признаки: необходимая пыльца для проведения гибридизации; уникальные и единичные семена, в том числе не выносящие обезвоживания; трансформированные, мутантные, гибридные клетки разных видов растений, способных к морфогенезу in vitro; зиготические и соматические зародыши и т.д. В настоящее время разработаны условия криосохранения для культивируемых клеток более 30 видов, каллусных культур (около 10 видов), изолированных протопластов (8 видов), сохранения меристем (25 видов) и кончиков стебля (13 видов). Приоритет в этом направлении принадлежит Институту физиологии растений РАН и, в частности, отделу культуры тканей и морфогенеза, возглавляемому проф. Р.Г. Бутенко.
При проведении работ по криосохранению необходимо, прежде всего, учитывать специфику растительных клеток: отбирать мелкие клетки, с маленькой вакуолью и пониженным содержанием воды; разрабатывать в каждом отдельном случае подходы замораживания и последующего оттаивания растительных клеток. При криосохранении встречается ряд трудностей, одна из которых связана с защитой замораживаемых клеток и тканей от осмотического стресса и механического разрушения структур в результате образования и роста кристаллов льда внутри клетки. Одновременно с этим необходимо правильно подбирать условия, обеспечивающие высокую выживаемость клеток при оттаивании и рекультивации.
Несмотря на многообразие работ в этом направлении, в них все же наметились общие приемы, лежащие в основе криосохранения: обработка клеток перед замораживанием, применение криопротекторов, соблюдение определенного режима замораживания в интервале от 0 до –40° С (в редких случаях до -70° С), а также специальные предосторожности при оттаивании объектов.
Процесс криоконсервации, как правило, начинается с подготовки культуры клеток к замораживанию. Это может быть достигнуто несколькими способами, предусматривающими культивирование клеток на питательных средах, содержащих различные осмотически активные вещества: маннит или сорбит в концентрации 2–6%, аминокислоты и среди них, в первую очередь, пролин, чье значение для связывания воды в клетках растений широко известно, а также у-аминомасляная кислота.
Подбор криопротекторов, веществ, уменьшающих повреждение клеток от осмотического и механического стресса, проводят эмпирически по принципу наименьшей токсичности и оптимального эффекта. Среди всех известных криопротекторов выделяются такие легко проникающие в клетки вещества, как диметилсульфоксид (ДМСО, 5–10%), глицерин (10–20%), а также непроникающие высокомолекулярные–поливинилпиролидон (ПВП), декстран, полиэтиленгликоль (ПЭГ) с молекулярной массой 6000.
Большое значение при криосохранении имеет правильно подобранный режим замораживания от 0 до –40° С. Как правило, для всех объектов устанавливается скорость замораживания 0,5–1 °С в минуту и всю эту работу проводят на специальном оборудовании, обеспечивающем программное замораживание. Такие приборы выпускает специальное конструкторское технологическое бюро с опытным производством при Институте проблем криобиологии и криомедицины (г. Харьков).
Таким образом, медленное замораживание и использование криопротекторов позволяет освободить клетку от свободной воды, и при –40° С клетки становятся полностью обезвоженными, что дает возможность проводить дальнейшее замораживание, а именно погружать ампулы с растительным материалом в жидкий азот.
Хранение материала в жидком азоте практически не лимитировано. Например, в криобанке Института физиологии растений РАН хранятся клетки моркови, которые находятся в жидком азоте около 20 лет, меристемы картофеля – более 10 лет и др.
Оттаивание и проверка жизнеспособности клеток после хранения в жидком азоте является последним этапом технологии криосохранения. Если замораживание осуществляют медленно, постепенно, то оттаивание должно быть проведено как можно быстрее. Для этого ампулы помещают в водяную баню с температурой 40°, а иногда и 60° С и выдерживают до полного исчезновения последнего кристаллика льда.
Для определения жизнеспособности клеток после оттаивания применяют наиболее простой, быстрый и вполне удовлетворительный способ – окраска витальным красителем (0,1%-ным феносафранином или 0,25%-ным раствором сини Эванса), в результате которой мертвые клетки окрашиваются, а живые нет. Окончательным критерием, безусловно, служит четкое возобновление роста и деления клеток при рекультивации на искусственных питательных средах после оттаивания.
Экспериментально было показано, что клетки после хранения в жидком азоте не теряют способности к делению, регенерации растений, не уменьшается продуктивность синтеза вторичных метаболитов (клетки продуценты) и т.д. Так, Институтом физиологии растений РАН совместно с НПО по картофелеводству разработаны методы криосохранения меристем четырех сортов картофеля и показана возможность из 20% хранящихся меристем регенерировать целые растения, которые при высадке в поле не отличались по всем признакам, включая темпы роста и продуктивность, от обычных пробирочных растений (С. Манжулин и др., 1982). Более подробно о технике криосохранения можно узнать из обзорных работ А.С. Попова.
Таким образом, технология, связанная с криосохранением растительных объектов, развивается и постоянно совершенствуется. Несомненно, эта технология имеет свое будущее, так как уже сегодня криобанки могут значительно облегчить работу селекционеров, предоставив им возможность широко использовать пул генов сортов, в том числе старой селекции и диких видов, а также исчезающих видов растений.
Похожие работы
... растением, количество веществ специализированного обмена. Только благодаря правильно разработанной стратегии получения высокопроизводительных штаммов к настоящему времени получены культуры тканей, в которых содержание вторичных продуктов достаточно велико, чтобы служить лекарственным сырьем. Однако для многих культур неоднократные попытки различных исследователей определить условия накопления ...
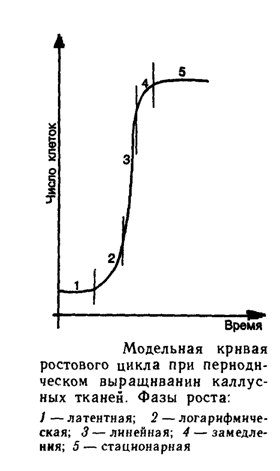

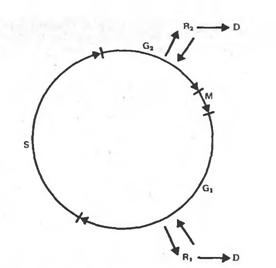
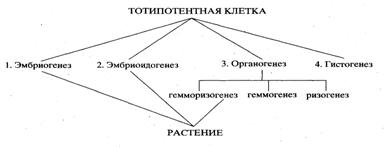
... по фосфору, азоту или сахару. Режим хемостата позволяет с помощью фиксированной скорости разбавления поддерживать константную скорость деления и плотность клеток в популяции. Клеточные суспензии в биотехнологии используются для получения вторичных метаболитов, многие из которых являются ценными лекарственными препаратами, для промышленного выращивания клеточной биомассы и для клеточной селекции. ...
... частности, биогенного происхождения: поли-b-оксибутирата, полиамилозы) и поиск новых активных микроорганизмов-разрушителей полимеров (полиэтилена, полипропилена, полихлорвинила). Усилия биотехнологов направлены также на борьбу с пестицидными загрязнениями — следствием неумеренного и нерационального применения ядохимикатов. Биотехнологические разработки играют важную роль в добыче и переработке ...
... на 54—71% и увеличивает урожай на 2 ц/га. Внесение триходермина в торфоперегнойные горшочки в количестве 50 мг снижает поражаемость огурцов корневыми гнилями на 60 % и увеличивает урожай на 31—74 %. Биотехнология и генетическая инженерия открывают новые широкие возможности в деле создания форм микроорганизмов, губительно влияющих на возбудителей болезней растений, отличающихся высокой ...
0 комментариев