Боль формируется как ответная реакция организма на разрушающие его раздражители. Данная реакция, по-видимому, выработалась в процессе эволюции живых существ как приспособительное свойство, направленное на улучшение выживаемости во внешней среде, так как при помощи болевого ощущения определялись те раздражители, которые непосредственно угрожают его жизнедеятельности или даже жизнеспособности. Если бы организм не обладал чувством боли, как это наблюдается в отдельных случаях у людей ("врожденная анальгия"), то своевременно определить вредоносность воздействия было бы невозможно, и смерть организма в таких условиях была бы более вероятной.
Можно предполагать, что боль является отрицательной биологической потребностью, формирующей перцептуально-мотивационный компонент той функциональной системы, которая контролирует два важнейших полезных приспособительных результата, две жизненно важные константы организма:
1. целостность его покровных оболочек, обеспечивающих изолированность от внешнего мира и тем самым постоянство внутренней среды организма,
2. уровень кислородного дыхания тканей, поддерживающий их нормальную жизнедеятельность.
Различают два основных вида боли – физическую и психогенную. В зависимости от причин физическую боль делят на три категории:
![]() Боль, обусловленная внешними воздействиями (за исключением боли от чрезмерных адекватных влияний на органы чувств, например, зрение, слух). Такая боль имеет следующие особенности: она всегда возникает на коже, непродолжительна, за исключением тех случаев, когда нарушается целостность кожи. Определить локализацию такой боли и установить причину ее возникновения легко. Возможно устранение внешнего воздействия. Нервная система остается неповрежденной , сохраняются целостность ее периферического аппарата и функций центральных механизмов, модулирующих болевое ощущение. В эту категорию входит большинство видов экспериментально вызванной боли. Боль, возникающая в результате внешних воздействий, может быть условным элементом в механизме обучения организма предупреждению повреждений. Эта категория включает боль, наблюдающуюся при клиническом сенсорном исследовании, которое определяет сохранность функций органов и тканей.
Боль, обусловленная внешними воздействиями (за исключением боли от чрезмерных адекватных влияний на органы чувств, например, зрение, слух). Такая боль имеет следующие особенности: она всегда возникает на коже, непродолжительна, за исключением тех случаев, когда нарушается целостность кожи. Определить локализацию такой боли и установить причину ее возникновения легко. Возможно устранение внешнего воздействия. Нервная система остается неповрежденной , сохраняются целостность ее периферического аппарата и функций центральных механизмов, модулирующих болевое ощущение. В эту категорию входит большинство видов экспериментально вызванной боли. Боль, возникающая в результате внешних воздействий, может быть условным элементом в механизме обучения организма предупреждению повреждений. Эта категория включает боль, наблюдающуюся при клиническом сенсорном исследовании, которое определяет сохранность функций органов и тканей.
![]() Боль, обусловленная внутренними процессами. При боли этой категории возбуждаются любые типы рецепторов и механизмы их активации могут быть различными. Возникающий при этом афферентный поток воспринимается как боль. В появлении такой боли кожа обычно не участвует, за исключением случаев ее прямого повреждения или отраженной боли. Локализация боли, идентификация ее причины пациентом, частичное или полное устранение источника боли нередко невозможны. Она имеет значительно большую длительность. При этой категории боли нервная система остается неповрежденной, так как фокус патологического процесса расположен дистальнее рецепторов. Кроме того, сохраняются нормальное поведение по афферентным волокнам и функции механизмов, модулирующих боль. По типу вовлекаемой ткани боль этой категории делят на эктодермальную, мезодермальную и эндодермальную, а также боль от чрезмерной нагрузки мышц.
Боль, обусловленная внутренними процессами. При боли этой категории возбуждаются любые типы рецепторов и механизмы их активации могут быть различными. Возникающий при этом афферентный поток воспринимается как боль. В появлении такой боли кожа обычно не участвует, за исключением случаев ее прямого повреждения или отраженной боли. Локализация боли, идентификация ее причины пациентом, частичное или полное устранение источника боли нередко невозможны. Она имеет значительно большую длительность. При этой категории боли нервная система остается неповрежденной, так как фокус патологического процесса расположен дистальнее рецепторов. Кроме того, сохраняются нормальное поведение по афферентным волокнам и функции механизмов, модулирующих боль. По типу вовлекаемой ткани боль этой категории делят на эктодермальную, мезодермальную и эндодермальную, а также боль от чрезмерной нагрузки мышц.
![]() Боль, связанная с повреждением нервной системы, в особенности ее афферентного аппарата. Хотя такая боль часто сопровождается ощущениями на коже, правильно идентифицировать внешние причины бывает затруднительно или невозможно. Также трудно определить локализацию источника боли. Боль этой категории продолжительная, может длиться годами и устранение ее источника невозможно. Нервная система повреждена: отмечаются дефекты проводящих периферических или центральных путей, а также нарушения функций механизмов, модулирующих боль. При боли этой категории повреждения лежат проксимальнее рецепторов в периферических нервах, спинном мозге или высших центрах мозга. Они могут быть локальными или системными (невралгии, каузалгии, фантомная боль, таламический синдром).
Боль, связанная с повреждением нервной системы, в особенности ее афферентного аппарата. Хотя такая боль часто сопровождается ощущениями на коже, правильно идентифицировать внешние причины бывает затруднительно или невозможно. Также трудно определить локализацию источника боли. Боль этой категории продолжительная, может длиться годами и устранение ее источника невозможно. Нервная система повреждена: отмечаются дефекты проводящих периферических или центральных путей, а также нарушения функций механизмов, модулирующих боль. При боли этой категории повреждения лежат проксимальнее рецепторов в периферических нервах, спинном мозге или высших центрах мозга. Они могут быть локальными или системными (невралгии, каузалгии, фантомная боль, таламический синдром).
В физической боли также различают:
§ Первичную боль (быструю, колющую острую),например, вызванную уколом иглы в кожу; она точно локализована, быстро исчезает после удаления стимула, не вызывает эмоциональной реакции;
§ Вторичную боль (медленную, невыносимую, жгучую), она появляется через 0,5 – 1 с после ощущения первичной боли, не имеет четкой локализации, остается некоторое время после удаления стимула, сопровождается изменениями функций сердечно-сосудистой и дыхательной систем, может влиять на характер личности, ее образ мышления (к вторичной боли относят также тупую боль в висцеральных органах и глубоких соматических структурах);
§ Хроническую боль (физическая боль, продолжающаяся длительное время у некоторых больных хроническими заболеваниями), для нее характерны сложные нервные механизмы эмоционального, аффективного и поведенческого уровня, выражающиеся в реактивной депрессии, которая делает человека недееспособным и в корне меняет его жизнь.
Психогенная боль связана с психологическими или социальными факторами, такими как эмоциональное состояние личности, окружающая ситуация, традиция культуры. Она имеет неопределенное начало, возникает без очевидной причины. Природа психогенной боли может быть неясна во многих отношениях. Часто отмечается расхождение между остротой боли, описываемой пациентом, и его поведением. Боль может не наблюдаться в ночные часы. Характерно плохо определяемое и изменяемое место боли, не всегда совпадающее с дерматомами или местом отраженной боли. Кроме того, психогенная боль несвязанна с внешними стимулами, может изменяться в зависимости от настроения. Она снимается действием антидепрессивных препаратов и других методов, уменьшающих эмоциональное напряжение.
Периферические системы боли
Рецепторы боли. По классификации A. Iggo (1977), рецепторы кожи делятся на механорецепторы, механоноцицепторные и ноцицепторы. Считается, что ноцицепторы представляют собой свободные нервные окончания немиелизированных волокон, образующие плексиморфные сплетения в тканях кожи, мышц и некоторых органов. Обнаружены и тонкие нервные волокна, связывающие их с рецепторами прикосновения, давления и температуры (волокна Тимофеева), что может служить основой возбуждения ноцицепторов при сильных сенсорных возбуждениях.
Если рассматривать ноцицепторы по механизму их возбуждения, то можно выделить два типа. Первый тип ноцицепторов – это механоцицепторов, так как их деполяризация происходит в результате механического смещения мембраны, что позволяет ионам натрия проникать внутрь клетки. Хотя они способны реагировать на введение ацетилхолина, которое блокируется холинолитиками, но при этом чувствительность рецептора к механическим раздражениям не снижается.
К механоцицепторам кожи относятся:
· Ноцицепторы кожи с афферентами А۵-волокон, возбуждающиеся механическими стимулами, почти не реагирующие на термические и совсем не реагирующие на химические раздражения. Рецептивные поля их узкие, повторные раздражения их инактивируют и у них быстро развивается адаптация (что характерно для первой эпикритической боли).
· Ноцицепторы эпидермиса с афферентами С-волокон, которые возбуждаются механическими стимулами, на нагревание или охлаждение не реагируют, имеют узкие рецептивные поля, легко адаптируются.
· Ноцицепторы мышц с афферентами А۵-волокон, расположенные на поверхности мышц и в местах перехода мышцы в сухожилие. Активируются особенно сильно при давлении тупыми предметами на укол иглой. Быстро адаптируются.
· Ноцицепторы суставов с афферентами А۵-волокон, которые возбуждаются только при чрезмерном сгибании или выкручивании суставов.
· Тепловые ноцицепторы кожи с афферентами А۵-волокон, которые возбуждаются на механические раздражения и нагревание 36-43°С и не реагируют на охлаждение.
Второй тип ноцицепторов – это хемоноцицепторы. Деполяризация их мембраны возникает при воздействии химических веществ (растворы хлористоводородной, серной и уксусной кислот, гистамин, ацетилхолин, окситриптамин, брадикинин и др.), т. е. тех веществ, которые в подавляющем большинстве нарушают окислительные процессы в тканях. Характерно, что после действия повреждающих раздражителей их чувствительность значительно повышается, и они приобретают способность реагировать на ранее не повреждающие механические и термические стимулы. Данный тип ноцицепторов локализуется как на покровных оболочках организма, так и в глубоких тканях, в том числе в висцеральных органах и особенно их много в оболочках кровеносных сосудов.
К хемоноцицепторам Л.Н. Смолин (1975) относит следующие:
· Подкожные ноцицепторы с афферентами С-волокон, активирующиеся механическими стимулами при сильном давлении на кожу и подкожном введении химических веществ.
· Ноцицепторы кожи с афферентами С-волокон, активирующиеся механическими стимулами и сильным нагреванием. Характерно, что при воздействии на кожу обезьян термального раздражителя от 36 до 43°С реагируют рецепторы с афферентами А۵-волокон, а от 41 до 53°С – ноцицепторы с афферентами С-волокон. Данные ноцицепторы медленно адаптируются.
· Ноцицепторы кожи с афферентами С-волокон, возбуждающиеся механическими стимулами и охлаждением до 15°С.
· Ноцицепторы мышц с афферентами С-волокон, активирующиеся на механические, термические и химические раздражители, в том числе на брадикинин и гистамин.
· Ноцицепторы внутренних паренхиматозных органов, локализующиеся, вероятно, главным образом в стенках артериол.
Таким образом, большинство механоцицепторов имеют афференты А۵-волокон, и они расположены так, что обеспечивают контроль целостности кожных покровов организма, суставных сумок, поверхности мышц.
Хемоноцицепторы расположены в более глубоких слоях кожи и передают импульсацию преимущественно через афференты С-волокон. Особенности локализации хемоноцицепторов наводят на мысль, что они контролирую дыхательные функции тканей, в том числе и покровных оболочек. По мнению P.W. Nathan (1976), болевые рецепторы делятся на реагирующие на само повреждение и на те, которые реагируют на результат этого разрушения.
Адекватные стимулы для боли. Пока не имеется определенного мнения о том, существуют ли адекватные стимулы для рецепторов боли. Высказан ряд гипотез, из которых наиболее распространена следующая: свободные нервные окончания в коже содержат одно или несколько химических соединений в пузырьках или гранулах. Эти соединения являются специфическими и выделяются в ответ на особую стимуляцию. Агент, высвобождающийся из нервных окончаний, взаимодействует с рецепторами наружной поверхности мембраны окончаний и вызывает ее деполяризацию. Действие соединения на рецепторы прекращается соответствующим ферментом, окружающим нервные терминали (см. рис. 1).
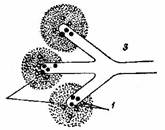
Рис. 1 Предполагаемая модель действия синаптического передатчика на свободное нервное окончание.
1 – рецепторная субстанция, 2 – разрушающий фермент, 3 – свободное нервное окончание.
В качестве активаторов рецепторов предположены ионы калия, гистамин, брадикинин, соматостатин, субстанция Р, простагландины. Однако ни одно из данных веществ еще не обнаружено в свободных нервных окончаниях. В вариантах этой гипотезы в качестве активаторов рецепторов свободных нервных окончаний предлагают рассматривать вещества протоплазмы клеток, разрушающихся при механических или термических воздействиях на ткань, а также продукты, образующиеся при воспалительном процессе. Полагают, что рецепторы терминалей активируются при локальной гипоксии участков тканей в ответ на механическое сдавливание капилляров вокруг свободных нервных окончаний. Высказана гипотеза о том, что изменение концентрации водородных ионов, приводящее к ацидозу в межклеточном пространстве, может обусловливать возбуждение рецепторов протонов, обнаруженных в мембране сенсорных нейронов ганглия тройничного нерва. Согласно другим предположениям, адекватным стимулом для рецепторов боли является сильная механическая деформация свободных нервных окончаний, вызванная сжатием ткани или перемещением жидкости в канальцах дентина зубов.
Предполагаемые рецепторные субстанции и синаптические передатчики. С помощью иммуногистохимического метода показано, что субстанция Р (пептид, обнаруженный во многих областях центральной нервной системы) распределяется вокруг терминалей чувствительных нервов. В непосрне5дственой близости от терминалей наблюдаются деградирующие ферменты – ацетилхолинэстераза (вокруг соматических окончаний) и бутирилхолинэстераза (вокруг висцеральных рецепторов).
Существует гипотеза о том, что каждый функциональный тип чувствительных нейронов спинномозговых узлов вырабатывает свою собственную специфическую рецепторную субстанцию и транспортирует ее аксоплазматическим током из тела клетки к периферическим окончаниям и, вероятно, к центральным окончаниям, где эта субстанция служит синаптическим передатчиком первичного афферентного волокна, т. е. рецепторная субстанция и синаптический передатчик сенсорного нейрона являются одним и тем же химическим веществом.
Одним из предполагаемых синаптических передатчиков ноцицептивных нейронов в заднем роге спинного мозга – субстанция Р, которую содержат 10-20% сенсорных нейронов спинномозговых узлов. Иммуногистохимические методы показали, что этот пептид локализуется в маргинальном и желатинозном слоях заднего рога, там, где оканчиваются волокна болевой чувствительности. Все нейроны в этих областях, которые активировались при стимуляции А-۵- и С-волокон, эффективно активировались также субстанцией Р.
Другим вероятным передатчиком сенсорных нейронов спинномозговых узлов считают пептид соматостатин, обнаруженный примерно в 10% этих нейронов. Установлено, что нейроны, содержащие субстанцию Р и соматостатин, относятся к популяции малых нейронов спинномозговых узлов, имеющих безмиелиновые аксоны.
По-видимому, периферические или центральные нейроны можно рассматривать как элементы системы боли, если эти нейроны отвечают исключительно на болевые стимулы, имеют анатомические связи с нейронами, образующими системы боли, если стимуляция этих нейронов вызывает ощущении боли и их ответы на болевой стимул уменьшаются одновременно со снижением ощущения боли.
Сенситизация. Общепризнано, что большинство типов сенсорных рецепторов становятся менее чувствительными, если подвергаются длительной стимуляции. В отличие от них у ноцицепторов чувствительность прогрессивно возрастает (до определенных пределов), если действие сохраняется продолжительное время. Такое снижение пороговой чувствительности называется сенситизацией. Это явление обнаружено также в процессе регенерации поврежденного нервного ствола, когда разрастающаяся соединительная ткань вызывает сжатие регенерирующих тонких нервных волокон. При этом в нерве регистрируется необычайно высокая фоновая ритмика и низкий порог для активации волокон в области повреждения. Согласно одной из гипотез о происхождении этого феномена, повреждение тканей вызывает высвобождение рецепторной субстанции, сопровождающееся инактивацией разрушающего ее фермента и стойкой деполяризацией мембраны рецептора, что обусловливает устойчивые разряды в афферентном волокне. Сенситизация проявляется в способности ноцицептора отвечать на адекватные стимулы допороговой величины (например, активироваться при температуре 40 °С, а не 45 °С, как отмечалось ранее), а также в способности возбуждаться ноцицептивными стимулами прежде неэффективных модальностей. Явление сенситизации многие исследователи рассматривают как одну из причин кожной гиперальгезии.
Центральные системы боли
Цитоархитектоника задних рогов спинного мозга. В дорсальных рогах спинного мозга находятся центральные нейроны, воспринимающие и перерабатывающие болевые сигналы перед отправкой их в головной мозг. Задний рог принято делить на 5 – 6 пластин. Первая пластина – краевая зона, содержит маргинальные нейроны, которые являются ноцицептивными интернейронами спиноталамического пути. Вторая – третья пластины образуют тесное сплетение мелких нейронов – желатинозную субстанцию, содержащую в себе малые тормозящие интернейроны, создающие сложные замкнутые цепочки, и крупные нейроны, аксоны которых идут в спиноталамический путь. Важной структурой желатинозной субстанции являются гломерулы – сложные синапсы между терминалями первичных афферентных волокон, дендритами центральных нейронов и окончаниями нисходящих путей. Эти структуры, как полагают, обеспечивают взаимодействие между афферентными и центральными потоками импульсов в процессе переработки информации. Четвертая и пятая пластины, или собственное ядро, - зона скопления самых крупных нейронов заднего рога, дендриты которых проникают в желатинозную субстанцию, а аксоны многих из них формируют спиноталамический путь. Нейроны собственного ядра легко активируются болевыми и неболевыми стимулами. Более глубокие пластины серого вещества (седьмая – восьмая), не относящиеся к заднему рогу, дают начало спиноретикулярным волокнам.
Центральные проекции первичных афферентных волокон. Перед входом в спинной мозг толстые и тонкие волокна дорсальных корешков перераспределяются. При этом тонкие миелиновые и немиелиновые волокна, несущие информацию о боли, группируются в отдельные пучки, которые входят в спинной мозг и образуют зону Лиссауэра. Пройдя в ней вдоль спинного мозга 1 – 2 сегмента, терминали волокон проникают в первую пластину серого вещества задних рогов. Толстые миелиновые волокна, войдя в спинной мозг, образуют петлю, входят в серое вещество с внутренней стороны задних рогов через пятую пластину и лишь затем достигают желатинозной субстанции (вторая – третья пластины).
Тонкие первичные афферентные волокна оканчиваются в первой половине – маргинальной зоне, а большинство толстых афферентных волокон – во второй пластине желатинозной субстанции. Перед окончанием толстые афферентные волокна проходят вдоль внутренней поверхности задних рогов, входят в серое вещество через четвертую – пятую пластины и затем поднимаются во вторую пластину. Здесь они оканчиваются факелоподобными пучками, но не проникают в маргинальный слой. Факелоподобные пучки окончаний толстых афферентных волокон делят желатинозную субстанцию на дольки, в которых нейроны организованы в вертикальные столбы. Крупные нейроны четвертой пластины посылают свои дендриты в эти дольки, где и устанавливаются синаптические контакты с толстыми первичными афферентными волокнами.
Считается доказанным, что нейроны второй – третьей пластин (область желатинозной субстанции) во взаимодействии с маргинальными нейронами образуют специализированную систему модулирующего контроля болевого афферентного входа, а пятая пластина является уровнем полимодального ноцицептивного входа и областью взаимодействия с толстыми неболевыми афферентами (см. рис. 2).
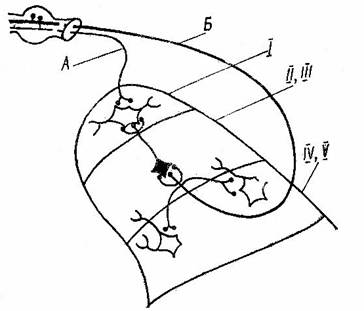
Рис. 2 Схема некоторых связей толстых и тонких первичных афферентов нейронами заднего рога.
А – тонкие, Б – толстые афференты, Й – маргинальный слой, ЙЙ и ЙЙЙ – желатинозная субстанция, ЙV и V – собственное ядро.
Тонкие афферентные волокна от кожи, мышц, сухожилий и внутренних органов (группы А-۵-, С, ЙЙ, ЙЙЙ, ЙV) через интернейроны заднего рога замыкается на мотонейронах сгибательных мышц и образуют обширную группу сегментарных сгибательных рефлексов, которые играют важную роль в ноцицептивных механизмах, создавая защитные рефлекторные реакции.
Функциональная организация нейронных цепей в заднем роге. В настоящее время предложены две основные схемы взаимодействия афферентных сигналов в заднем роге. Вопрос о входе к маргинальным нейронам пока не решен. Зона Лиссауэра содержит не только первичные афферентные волокна, но и аксоны нейронов желатинозной субстанции. После перерезки этой зоны аксосоматические и аксодендритные синапсы на маргинальных нейронах дегенерируют, что указывает на то, что С-волокна и аксоны нейронов желатинозной субстанции должны оканчиваться на них. Как уже упоминалось, С-волокна оканчиваются в маргинальной зоне и желатинозной субстанции. В свою очередь, нейроны желатинозной субстанции образуют тормозящие синапсы на соме маргинальных нейронов.
Толстые афферентные волокна заканчиваются возбуждающими синапсами на нейронах желатинозной субстанции и больших нейронах собственного ядра. Полагают, что ритмическая активность нейронов желатинозной субстанции импульсами от толстых афферентных волокон может выражаться в интенсивном торможении маргинальных нейронов, на которых образуют свои синапсы тонкие волокна. Такая нейронная цепь может создать механизм, посредством которого толстые неболевые афферентные волокна моделируют порог боли. На рисунке показана принципиальная схема возможной организации такой системы контроля.

Рис.3 Схема контроля интенсивности сенсорного потока в заднем роге серого вещества спинного мозга.
А – тонкие, Б – толстые афференты, ЖС – желатинозная субстанция, Т – нейроны, передающие сенсорную информацию в мозге, (+) – возбуждение, (-) – торможение.
Толстые неболевые афференты и тонкие болевые афференты подходят к нейронам в желатинозной субстанции и центральным нейронам, передающим сенсорную информацию в мозг (Т-клетки). Тормозящий эффект от нейронов желатинозной субстанции, возникающий в аксоаксональных синапсах на толстых и тонких афферентах, увеличивается, если возрастает поток импульсации в толстых неболевых афферентах, и уменьшается, если повышается поток импульсации по тонким болевым афферентам. Существуют также механизмы центрального контроля, которые управляются потоком импульсации по толстым афферентам. Эти механизмы проецируются в воротную систему контроля, Т-клетки посылают сигналы в систему действия.
Известно, например, что боль, возникшая при порезе пальца, уменьшается при давлении на окружающие структуры. По-видимому, повышение активности в толстых волокнах уменьшает передачу сигнала от тонких волокон. При использовании местных раздражающих средств снижение боли объясняется подобным образом: высокочастотные разряды, возникающие в терморецепторах в ответ на местное тепловое раздражение, уменьшают передачу импульсов от тонких ноцицептивных волокон и таким образом уменьшают ощущение боли.
Нейроны серого вещества спинного мозга, как полагают, не образуют единую восходящую систему боли. Вероятно, поэтому 50% лиц, которым проводилась частичная перевязка проводящих путей спинного мозга, прерывающая главные восходящие пути (антеролатеральная хордотомия), в связи с непереносимой болью, отмечалось повторное появление боли.
Всходящие системы, передающие ноцицептивную информацию
По-видимому, в мозге нет узкоспециализированных восходящих систем, передающих только болевую информацию. Однако в ряде из них объем ноцицептивных потоков достаточно велик. К таким системам относятся спиноталамические, спиноретикулярные, спиномезэнцефалические и спиноцервикальные пути, а также проприоспинальная система спинного мозга и проприоретикулярная система ствола мозга.
Установлено, что многие нейроны каждой из пластин заднего рога посылают импульсы к клеткам более глубоких слоев, которые формируют длинные восходящие пути, однако большинство нейронов, дающих длинные восходящие аксоны, обнаружены в первой, второй и пятой пластинах. Существовавшее долгое время мнение о том, что спиноталамический путь служит исключительно для передачи болевой информации, в настоящее время пересмотрено. Сейчас доказано, что боль является не единственной модальностью, передающейся в таламус этим путем. Спиноталамический путь начинается от нейронов, которые лежат преимущественно в первой и пятой пластинах заднего рога. Многие из них проецируются на контралатеральную сторону спинного мозга. Большая часть этих нейронов возбуждается как неболевыми, так и болевыми стимулами. Спиноталамический путь делится на неоспино- и палеоспиноталамический путь.
Неоспиноталамический путь, или латеральный, путь – филогенетически молодая система длинных волокон, лежащая в латеральных отделах спинного мозга и ствола мозга, устанавливает контакты с теми нейронами вентролатерального и заднего таламуса, которые посылают аксоны в первую соматосенсорную область коры большого мозга. Соматотопически четко организованный, этот путь осуществляет передачу тактильной и болевой информации о положении периферического стимула в пространстве и времени, его интенсивности и продолжительности. Через него передается информация об острой боли, вызванной повреждением тканей.
Палеоспиноталамический, или вентральный, путь лежит в медиальной части ствола мозга и распределяет свои окончания в медиальных и интраламинарных ядрах таламуса, гипоталамуса, структурах лимбического мозга. Его функции охватывают недискриминативные аспекты боли – через этот путь передается тупая, слаболокализованная боль. Палеоспиноталамический путь приспособлен для формирования разнообразных супрасегментарных рефлекторных ответов, связанных с дыханием, кровообращением, эндокринными функциями, через него запускаются мощные мотивационные и защитные реакции.
Спиноретикулярный путь является важным путем передачи болевой информации. Он берет начало от нейронов глубоких пластин заднего рога, а также от серого вещества переднего рога спинного мозга. Окончания этих нейронов распределяются в ядрах ретикулярной формации продолговатого мозга и моста, главным образом в ретикулярном гигантоклеточном ядре, которое представляет собой основное передаточное ядро болевой информации в спиноретикулярной системе. Нейроны этого ядра возбуждаются при естественной болевой стимуляции и электрическом раздражении тонких миелиновых волокон. Неясно, однако, важны ли сигналы, идущие по этому пути, для сознательного восприятия боли или они вызывают только рефлекторные ответы на боль. Обширные связи ретикулярной формации с гипоталамусом и лимбическими структурами дают основание предполагать, что ретикулярная формация играет важную роль в мотивационных и аффективных состояниях, связанных с болью.
Спиномезэнцефалический путь начинается от тех же областей серого вещества спинного мозга, что и Спиноретикулярный путь. Его нейроны распределяют свои окончания в вентромедиальной ретикулярной формации среднего мозга. Нейроны этого пути имеют широкие рецептивные поля и легко активируются болевыми стимулами различного происхождения. Функциональная организация спиномезэнцефалического пути изучена мало. Полагают, что болевая информация, поступающая по нему, способствует приведению организма в готовность выполнять защитные реакции.
Спиноцервикальный путь формируется из аксонов нейронов четвертой – седьмой пластин серого вещества спинного мозга. Его нейроны активируются как болевыми (термическими, механическими и химическими), так и неболевыми стимулами, аксоны этого тракта поднимаются в заднебоковом спинальном канатике к латеральному шейному ядру спинного мозга. Спиноцервикальный путь хорошо развит у грызунов, хищников, слабо – у приматов и человека.
Проприоспинальная система спинного мозга образована тремя подсистемами: дорсолатеральной проприоспинальной системой, дорсальным интракорнуальным трактом и латеральной частью зоны Лиссауэра. Общим для этих подсистем является объединение в бесчисленные цепочки множества короткоаксоновых интернейронов, тела которых лежат в первой и пятой пластинах серого вещества, а аксоны распространяются в пределах нескольких сегментов спинного мозга. Аналогично организованы и проприоретикулярные системы, объединяющие лежащие наиболее медиально ретикулярные структуры спинного, продолговатого, среднего мозга и моста. Такие мультисинаптические восходящие системы создают дополнительные и сложные пути для передачи тупой слаболокализованной боли. Возможно, что поступающая ноцицептивная информация прежде чем достичь длинных восходящих систем, некоторое время может передаваться через короткие межнейронные связи. Этим объясняют, почему уровень анальгезии, вызванной вентролатеральной трактотомией (перерезкой проводящих путей спинного мозга), начинается на несколько сегментов каудальнее перерезки.
Система «задние столбы – медиальная петля» также играет важную роль в передаче боли. Клинические наблюдения показывают, что повреждение задних столбов спинного мозга у человека приводит к глубокой анестезии, однако их стимуляция может уменьшать чувство боли – метод, используемый для снижения боли у лиц с хроническими заболеваниями. В эксперименте показано, что во многих волокнах задних столбов обнаруживают ответы на болевые механические и термические стимулы, а увеличение активности в волокнах заднего столба обусловливает подавление передачи болевой импульсации через системы задних рогов спинного мозга. Имеются сведения о том, что импульсация по волокнам задних столбов запускает активность систем головного мозга, участвующих в пространственном и временном анализе качеств болевого стимула, идущего через другие входы. Благодаря быстрой передаче информации об этих качествах, высшие отделы мозга успевают идентифицировать, оценивать, локализовать и избирательно модулировать через быстрые нисходящие пути сенсорный вход перед тем, как будет включена система действия.
Переработка восходящей ноцицептивной информации в центральных структурах мозга. Экспериментальные и клинические наблюдения указывают на то, что боль как модальность обеспечивается пространственно распределенной системой центральных структур. Нейроны, связанные с ноцицепцией, обнаруживаются, например, в спинном мозге, ретикулярной формации, центральном сером веществе (ЦСВ), верхних бугорках крыши среднего мозга, латеральном и медиальном таламусе, гипоталамусе, лимбической системе, различных областях коры большого мозга и некоторых других структурах и в ответ на ноцицептивную стимуляцию создают присущие только ноцицептивной системе и качественно отличающиеся компоненты общей реакции организма на боль.
С помощью нейрофизиологических исследований, а также наблюдений за поведением человека и животных с повреждением или разрушением или раздражением участков мозга установлено, что через спинной мозг реализуются моторные и симпатические рефлексы, там же происходит первичная обработка ноцицептивных сигналов; ретикулярной формацией ствола мозга контролируется кровообращение и дыхание; гипоталамус поддерживает гомеостазис и регулирует выделение гормонов из гипофиза; базальные ганглии и мозжечок осуществляют двигательное программирование; через лимбическую систему реализуются аффективно-мотивационные, а через кору большого мозга – когнитивные компоненты болевого поведения.
Анализ организации нейронных цепей в задних рогах спинного мозга позволили сформулировать концепцию о том, что спинной мозг представляет собой интегративный центр для сенсорных процессов, а не передаточный механизм. В спинном мозге отдельные сенсорные модальности не передаются по жестко определенным восходящим путям, а подвергаются сложной переработке. В его многочисленных нейронных цепях и синаптических контактах происходят различные интегративные процессы, приводящие к тому, что значительное число импульсов, входящих в спинной мозг, отфильтровывается и не достигает высших отделов мозга. Системы фильтрации афферентных входов в спинном мозге гибкие и легко перестраиваются. Появление болевого сигнала может полностью отключить другие сенсорные входы до того момента, пока опасность не будет преодолена. И наоборот, в отдельных ситуациях интенсивный неболевой вход или волевое усилие могут подавить двигательные рефлекторные движения, вызванные болевыми стимулами.
Ретикулярная формация занимает важное положение в головном мозге, простираясь от спинного мозга до границы между средним и промежуточным мозгом. В ней заканчиваются или к ней отдают коллатерали длинные восходящие пути, несущие разнообразную сенсорную, в том числе и ноцицептивную информацию. Ретикулярная формация располагает мощной проприоретикулярной системой, тесно взаимосвязана с различными сенсорными и вегетативными системами и постоянно контролирует двигательные системы ствола мозга и спинного мозга. Здесь обнаружены ретикулярные нейроны, активирующиеся широким спектром афферентных потоков различных модальностей и реагирующие исключительно на ноцицептивные стимулы. Благодаря этому ретикулярная формация выполняет широкие функции по переработке ноцицептивной информации. К ним относятся: подготовка и передача ноцицептивной информации в высшие соматические и вегетативные отделы мозга (таламус, гипоталамус, лимбическую систему, кору); усиление и пролонгирование защитных сегментарных рефлексов спинного мозга и ствола мозга; вовлечение и рефлекторный ответ на ноцицептивные стимулы вегетативной нервной системы, прежде всего симпатической (увеличение частоты сердечных сокращений, подъем артериального давления и т. д.); включение сложных рефлекторных ответов структур ствола мозга, сопровождающих защитные реакции организма и приводящих к пробуждению, настораживанию организма и его готовности отвечать на потенциально опасные ситуации.
Верхние бугорки крыши среднего мозга – один из нервных центров сенсомоторной интеграции, специфически связанный со зрительным поведением – участвуют в переработке ноцицептивных потоков, облегчающих зрительную ориентацию в момент действия болевых стимулов.
ЦСВ среднего мозга и ядра шва продолговатого мозга являются звеньями системы подавления боли. Активация этих нейронов приводит к выделению нейропептидов (энкефалинов, эндорфинов) и моноаминов (серотонина, норадреналина), угнетающих передачу нервных импульсов от сенсорных нейронов на первые переключающие нейроны центральной нервной системы в местах распределения терминалей тонких, безмиелиновых афферентных нейронов, несущих болевую информацию (задние рога серого вещества спинного мозга, сенсорные ядра тройничного нерва).
Структуры латерального таламуса обеспечивают анализ сенсорно-дискриминативных качеств боли (соматотопика болевого стимула, его длительность, интенсивность и т. д.), тогда как медиальные таламические области участвуют в мотивационно-аффективных аспектах боли (быстрое выделение вновь появившихся соместетических стимулов, быстрая передача информации о проявлении новой потенциально опасной ситуации).
Ноцицептивная информация, поступающая в структуры гипоталамуса, активирует его нейронные и нейрогормональную системы. Это сопровождается развитием сложного комплекса длительных вегетативных и соматических реакций, направленных на перестройку мышечной и висцеральной систем в условиях продолжающегося действия болевых стимулов. Выброс в кровеносное русло гипоталамо-гипофизарных гормонов вазопрессина и окситоцина, помимо их участия в сложных поведенческих эффектах, по-видимому, вызывает депрессивное действие на сенсорные входы, повышая порог чувствительности этих входов.
Лимбическая система играет важную роль в сенсорных процессах боли, участвуя в создании эмоциональной окраски поведения организма при выполнении реакций защиты, бегства и нападения в ответ на ноцицептивную стимуляцию.
Хотя имеются многочисленные доказательства тесных взаимосвязей между ядрами таламуса и различными областями коры большого мозга, однако оценка функций коры большого мозга в переработке болевой информации противоречива. Полагают, что область первичного восприятия соматосенсорной информации (постцентральная извилина) не существенна для восприятия боли. Имеются указания на то, что этот процесс происходит в зонах орбитальной и перфронтальной коры большого мозга. В любом случае, поступление импульсов в кору большого мозга создает условия для возбуждения пирамидной и экстрапирамидных нисходящих систем, вызывающих комплекс последовательных сокращений мышц, необходимых для осуществления поведенческих реакций.
Однако, по данным ряда наблюдений, первичная и вторичная воспринимающая области коры большого мозга влияют на «осознание» болевых воздействий: при повреждении этих областей у людей отмечались анальгезия или гиперальгезия, а также спонтанно возникающие ощущения боли или состояния гиперпатии. Нередко удаление раздражающих кору большого мозга локальных повреждений или хирургические разрушения ее супракаллозальных областей приводит к прекращению упорной хронической боли у многих пациентов.
Таким образом, данные об анатомических путях, физиологические доказательства принципов организации нейронных систем и клинико-физиологические описания субъективных ощущений боли показывают, что афферентная система боли состоит из двух подсистем, выполняющих различные функции. Первая подсистема, начинающаяся быстро проводящими миелиновыми А-۵-волокнами, является предостерегающей системой. Ее основная задачам – обеспечить мозг немедленной информацией о наличии повреждения, его степени и точной локализации. Вторая подсистема с тонкими безмиелиновыми С-волокнами действует как напоминающая система. Генерируя особенно неприятную, медленную, диффузную, постоянную боль, вторая система постоянно напоминает мозгу о том, что повреждение наступило и, следовательно, должна быть ограничена нормальная активность и уделено внимание повреждению. Классическое различие между первичной и вторичной болью отражает различие между предостерегающей и напоминающей системами.
Нисходящие системы, участвующие в ноцицепции. Значительная часть структур мозга приспособлена для отбора, модуляции и контроля восходящей сенсорной информации через нисходящие системы. Некоторые из этих нисходящих систем исключительно важны для модуляции и контроля ноцицептивной информации. К таким структурам относятся кора большого мозга, ЦСВ и ядра шва ствола мозга. Импульс, возникающие в нейронах этих структур, могут не только влиять на передачу от сенсорных волокон на уровне первых синапсов в заднем роге спинного мозга, но и действовать на уровне ствола мозга и промежуточного мозга, модулируя передачу ноцицептивной информации. Принято считать, что каждая структура мозга, получающая восходящую сенсорную информацию, имеет нейроны с нисходящими аксонами, которые способны изменять объем сенсорного потока, поступающих в эту структуру.
Компоненты боли
Подытоживая все выше сказанное, можно выделить в общей системной болевой реакции несколько ее относительно самостоятельных компонентов
Ø Перцептуальный компонент – собственно ощущение боли, возникающее на основе возбуждения механо- и хемоноцицепторов. Это формирует ноцицептивную импульсацию, которая через афферентные пути активирует лемнисковые и экстралемнисковые системы спинного и головного мозга (вплоть до коры больших полушарий), создавая ощущение боли.
Ø Рефлекторная защитная двигательная реакция устранения от вредоносного фактора, формирующаяся на основе активации мотонейронов спинного мозга через конвергенцию на них ноцицептивной импульсации.
Ø Болевое "arousal", связанное с активацией ретикулярной формации, с последующими восходящими и нисходящими активирующими влияниями на другие структуры мозга и функциональные системы организма.
Ø Отрицательная эмоция, формирующаяся на основе возбуждения отрицательных эмоциогенных зон гипоталамуса, ретикулярной формации и лимбических структур, имеющая, по-видимому, преимущественно адренергическую природу, и вызывающая совместно с ноцицептивной импульсацией изменения вегетативных реакций организма, гормональных и метаболических процессов.
Ø Мотивация устранения болевых ощущений, формируется на основе активации лобных и теменных областей коры и приводящая формированию поведения, направленного на лечение ран или выключения перецептуального компонента.
Ø Активация механизмов памяти, связанная с извлечением опыта по устранению болевых ощущений, т. е. избегания вредоносного фактора или сведения до минимума его действия и опыта лечения ран.
Таким образом, болевая реакция «является интегративной функцией организма, которая мобилизует самые разнообразные функциональные системы для защиты организма от воздействующих вредящих факторов и включает такие компоненты, как сознание, ощущения, память, мотивации, вегетативные, соматические и поведенческие реакции, эмоции» (Анохин П.К., Орлов И.В., 1976).
Теории боли
В середине XX в. Эволюция идей о боли развивалась по нескольким концептуальным направлениям. Был выдвинут ряд теорий, которые подвергались острой критике по мере накопления фактов по их основным положениям. В 1965 г. R.Melzack, P. Wall предложили теорию «входных ворот», которая, несмотря на определенные недостатки, в настоящее время принята большинством исследователей. Авторы теории сделали попытку объяснить с позиций современных нейрофизиологии, нейроанатомии, психологии и клиники боли высокую специализацию рецепторов и сенсорных проводящих путей в мозге, роль временной и пространственной суммации в функциях нервных структур, участвующих в процессе восприятия и переработки информации о боли, влияние различных психологических процессов на восприятие боли и формирование ответа на нее, различные клинические проявления острой и хронической боли.
Основные положения этой теории:
ü Передача нервных импульсов от афферентных волокон к нейронам спинного мозга, передающим сигналы в головной мозг, модулируется «спинальным воротным механизмом» - системой интернейронов, локализованных в желатинозной субстанции (вторая и третья пластины заднего рога) спинного мозга.
ü Спинальный воротный механизм регулируется относительным количеством импульсов, поступающих по афферентным волокнам большого и малого диаметра.
ü Спинальный воротный механизм находится под влиянием нервных импульсов, которые передаются по волокнам нисходящих систем из коры большого мозга и ствола мозга.
ü Специализированная система волокон большого диаметра, обладающая высокой скоростью проведения, является триггером (пусковым устройством) центрального контроля, который активирует избирательные когнитивные процессы, влияющие через нисходящие волокна на модилирующие свойства спинального воротного механизма.
ü Когда возбуждение релейных нейронов спинного мозга превышает критический уровень, то их импульсация приводит к возбуждению системы действия.
Таким образом, основные принципы теории «входных ворот» учитывают специфичность рецепторов, физиологические механизмы конвергенции, суммации, облегчения и торможения, роль восходящих и нисходящих систем головного и спинного мозга. Тем не менее, ряд экспериментальных данных, связанных с эффектами действия тонких, болевых афферентов на передачу информации в головной мозг, не укладывается в различные варианты схем воротной системы. Теория «входных ворот» постоянно усовершенствуется с получением новых экспериментальных данных.
Мысль о том, что боль является состоянием, противоположным состоянию удовольствия, была широко распространена до начала XIX в. Появившиеся затем новые теории боли, основанные на экспериментальных данных, вытеснили эту точку зрения, но, как оказалось, лишь на время. Изучая развитие и течение боли, наблюдаемой при повреждении органов, что боль слабо связана с повреждением, но существенно – с состоянием организма. В связи с этим в состоянии организма после получения им повреждения автор выделяет три фазы:
o ближайшую (боль иногда не возникает совсем, но обязательно отмечаются некоторые формы активности: борьба, защита, поиски безопасного места и помощи);
o острую (возникает после того, как организм обезопасит себя от источника повреждения, формируется поведение организма, которое позволяет начать выздоровление, есть боль, сигнализирующая о необходимости начала такого поведения);
o хроническую (полное отсутствие активности, что является оптимальным условием выздоровления, однако сопровождается болью и депрессией и нередко длится дольше необходимого для выздоровления период.
Отрицая роль боли как полезной предохранительной меры, P. Wall предлагает считать ее сигналом о необходимости перехода организма в состояние отдыха и восстановления. Это ставит, по его мнению, боль в класс тех общих чувств (жажда, голод), которые называются мотивационными состояниями и которые побуждают организм устранять причины, вызывающие общие чувства.
В XX в. Наука о боли обогатилась важными концептуальными представлениями. Были сформулированы понятия острой и хронической, нормальной и патологической боли. Так, с точки зрения физиологов, острая боль – это боль короткой продолжительности, которая может быть вызвана и прекращена действиями экспериментатора (электрический стимул, радиационное тепло). Термин «хроническая боль» отражает болевую стимуляцию, которая длится часы, дни и даже недели, но которая все еще находится под контролем экспериментатора. Она может быть результатом стимуляции (ритмического раздражения через вживленные электроды), экспериментально вызванного повреждения или создания специальных условий (невромы, обусловленной перевязыванием и инкапсуляцией нерва). Для клиницистов и физиологов, использующих в работе адекватные натуральные болевые стимулы, «острая боль» может быть определена как проходящая боль, которая сигнализирует о повреждении ткани, или боль, связанная с ощущением повреждения ткани. Такая боль обычно уменьшается во время естественного процесса заживления. Если боль повторяется или имеет циклическое течение, например, при стенокардии, мигрени, болевых тиках, то ее называют возвратной. Хроническая или постоянная боль может быть определена как боль, которая не исчезла после окончания процесса выздоровления (после заболевания или повреждения). Сюда может быть отнесена боль, связанная с прогрессирующим злокачественным или незлокачественным заболеванием, наблюдающимся продолжительный период, при котором выздоровление не наступает, например, боль, связанная с раком или артритом.
В понятиях «нормативная» и «патологическая» боль причина боли рассматривается под иным углом. Нормальная боль возникает в тех случаях, когда происходит активация ноцицептивных афферентных окончаний от адекватных болевых стимулов. К патологической боли авторы относят боль, вызванную другими факторами, например, генерацией эктотопических импульсов в поврежденном нерве, ненормальными эпилептиформными разрядами в структурах центральной нервной системы, угнетением действующих в нормальных условиях нервных механизмов, подавляющих передачу болевых сигналов. При этом нормальная боль рассматривается как адекватная функция нервной системы, предназначенная для предупреждения о потенциально опасных для организма ситуациях, а патологическая боль – как проявление повреждения или заболевания нервной системы. Она часто продолжается долго после того, как исчезли признаки нарушения тканей, и может представлять собой более серьезную причину для беспокойства, чем повреждения или заболевания, которые ее вызвали.
Похожие работы
... рассуждения и клинические данные указывают на необходимость исследования анальгетических свойств препаратов, которые, судя по их химической структуре, могут влиять на аномальную экспрессию натриевых каналов в патологических условиях. 1.2 Анализаторы боли (на примере зубной и головной) Взаимодействие и взаимовлияние сна и головной боли двойственны: с одной стороны, сон может быть провокатором ...
... , достигаемой терморегуляцией. Температура кожи ниже внутренней температуры тела (примерно З6,6°С) и различна для отдельных участков (на лбу 34-35, на лице 20-25, на животе 34, на стопах ног 25-27°С). В коже человека находятся два вида анализаторов температуры: одни реагируют только на холод, другие - только на тепло. Всего на коже около 30 тысяч тепловых точек и примерно 250 тысяч точек холода. ...
... и утрачивается к началу полового созревания. 36. Учение Павлова о типах ВНД. Общин типы ВНД. Пластичность ВНД. Учет типов ВНД в индивидуальном обучении и воспитании Среди вопросов физиологии ВНД человека особое значение для совершенствования учебно-воспитательного процесса и разработки естественнонаучных теорий воспитания и обучения имеет учение о типах ВНД. Это связано с тем, что именно ...



... извилине (в области проекции лица). Афферентные связи вестибулярного аппарата. Г - глаз; М - мышца; Ж - желудок; ТК - тонкая кишка; ПМ - продолговатый мозг; CM - спинной мозг. Висцеральный анализатор. интерорецепторы Рецепторы, расположенные в органах, называются висцерорецепторами, или интерорецепторами. Импульсация от интерорецепторов поступает в продолговатый мозг по волокнам IX ( ...
0 комментариев